Effective strains enable rapid wound closure in jellyfish after injury
Pith reviewed 2026-05-21 06:16 UTC · model grok-4.3
The pith
Radially contractile pre-strains in jellyfish tissue enable mechanical wound closure even after large cuts.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
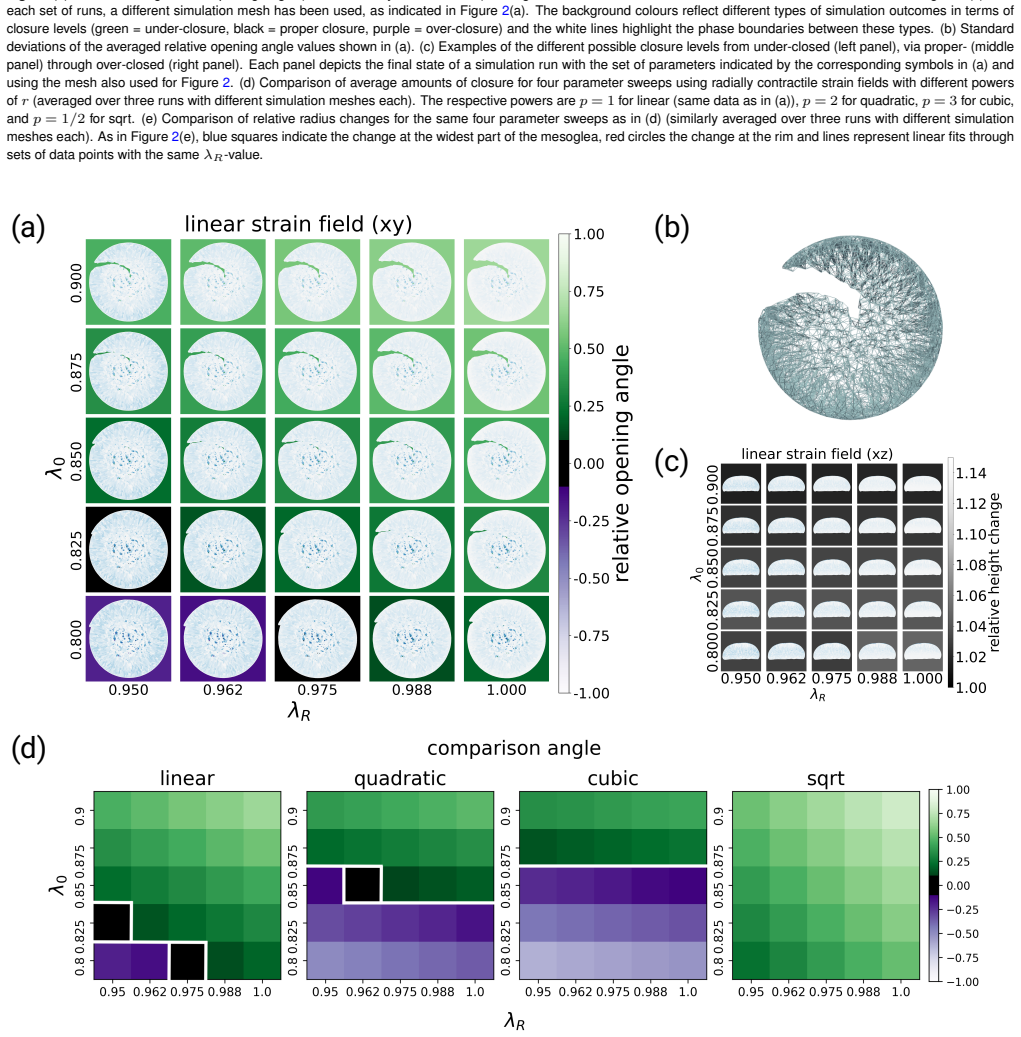
Using an in silico spring lattice model of the jellyfish umbrella, radially contractile pre-strains initiate wound closure over a wide range of conditions, including large cuts that leave only small pieces of tissue. This holds in agreement with experimental findings. An analytical expression gives the expected closure as a function of the residual material angle.
What carries the argument
Spring lattice model of the jellyfish umbrella geometry with imposed radially contractile pre-strains.
If this is right
- Wound closure proceeds mechanically for large injuries with minimal remaining tissue.
- Only radially contractile strains succeed; radially extensile strains do not.
- The fraction of closure is predictable from the residual material angle via a closed-form expression.
- Pre-strains suffice to explain the rapid pace seen in experiments.
Where Pith is reading between the lines
- Similar pre-strain mechanisms may operate in other fast-regenerating organisms.
- Tissue engineering could use controlled pre-strains to accelerate healing without relying on cell growth.
- Biological control of radial contractility in the jellyfish could be modeled as the source of the effective strains.
Load-bearing premise
The spring lattice model captures the essential mechanical behavior of the tissue so that pre-strains alone can produce the observed closure without other biological contributions.
What would settle it
Experiments that directly measure tissue strains during closure and find no significant radial contraction, or that show closure fails when contractile elements are disrupted while proliferation remains possible.
Figures






read the original abstract
The jellyfish Clytia hemisphaerica possesses astounding regenerative capacities and is able to close even large wounds within a few hours. This rapid pace of wound closure raises the question whether tissue mechanics, rather than tissue restructuring or cell proliferation, might be underlying the process. We tested this possibility by asking if simple pre-strains within the jellyfish umbrella would be capable of initiating wound closure in a jellyfish body geometry. To this end, we employed an in silico spring lattice model, a coarse-grained model of elastic materials which has previously been established to study tissue mechanics problems. We found that, using radially contractile (but not radially extensile) strains, wound closure can indeed be initiated across a wide range of conditions. This is even true for large cut sizes and, hence, small pieces of remaining tissue material, in good agreement with the experimental findings. Finally, we derived an analytical expression for the expected amount of achievable closure as a function of the residual material angle. These results establish important foundations for further investigations of the biophysics underpinning jellyfish regeneration.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper claims that radially contractile pre-strains applied within an established spring lattice model of the jellyfish umbrella geometry are sufficient to initiate and drive wound closure, even for large cuts leaving only small residual tissue angles, in agreement with experimental observations on Clytia hemisphaerica. An analytical expression is also derived for the expected closure as a function of the residual material angle.
Significance. If the model parameters faithfully represent the passive mechanics of the umbrella tissue, the result would demonstrate that pre-existing contractile strains alone can explain rapid wound closure without invoking cell proliferation or active restructuring, providing a mechanical foundation for jellyfish regeneration studies and a falsifiable prediction via the analytical closure formula.
major comments (2)
- [Methods (spring lattice model implementation)] Methods section on the spring lattice model: no calibration or direct comparison is reported between the chosen spring stiffnesses, connectivity, or pre-strain magnitudes and measured force-extension curves, bending stiffness, or anisotropy of Clytia hemisphaerica mesoglea plus epithelia. Because the central claim is that contractile (but not extensile) strains suffice for closure even at small residual angles, the absence of such validation leaves open the possibility that the simulated behavior is an artifact of untested parameter choices rather than a robust mechanical prediction.
- [Results (wound closure for large cuts)] Results on large-cut simulations: the reported closure for small remaining tissue angles relies on the specific pre-strain implementation and geometry; without a sensitivity analysis to variations in Poisson ratio, thickness, or viscous damping (none of which are stated to have been performed), it is unclear whether the agreement with experiments holds under plausible changes to the tissue constitutive response.
minor comments (2)
- [Abstract] The abstract states 'good agreement with the experimental findings' without specifying the quantitative metrics (e.g., closure time, final area, or shape) used for comparison; adding this would strengthen the claim.
- [Analytical derivation] Notation for the residual material angle and the derived analytical expression should be introduced with a clear equation number and diagram reference to aid readability.
Simulated Author's Rebuttal
We thank the referee for their thoughtful and constructive comments, which have helped us improve the clarity and robustness of the manuscript. We address each major comment in detail below and have revised the manuscript where appropriate to incorporate additional discussion and analyses.
read point-by-point responses
-
Referee: [Methods (spring lattice model implementation)] Methods section on the spring lattice model: no calibration or direct comparison is reported between the chosen spring stiffnesses, connectivity, or pre-strain magnitudes and measured force-extension curves, bending stiffness, or anisotropy of Clytia hemisphaerica mesoglea plus epithelia. Because the central claim is that contractile (but not extensile) strains suffice for closure even at small residual angles, the absence of such validation leaves open the possibility that the simulated behavior is an artifact of untested parameter choices rather than a robust mechanical prediction.
Authors: We agree that explicit calibration against Clytia-specific mechanical data would strengthen the presentation. The spring lattice model is a previously established coarse-grained framework for elastic tissue mechanics, and our parameter choices (stiffness, connectivity, and pre-strain magnitude) were selected to be consistent with typical values reported for soft biological tissues in the literature. The central qualitative result—that radially contractile pre-strains produce closure while radially extensile pre-strains do not—arises from the directional geometry of the umbrella and the sign of the strain rather than from precise numerical values of the spring constants. The analytical closure formula we derive is obtained from a purely geometric argument on the residual material angle and is therefore independent of the discrete-model parameters. In the revised manuscript we have expanded the Methods section to include a dedicated paragraph on parameter selection, with references to prior validations of the same modeling approach, and we have added a brief discussion of the expected range of pre-strain magnitudes based on known contractile properties of epithelial tissues. revision: partial
-
Referee: [Results (wound closure for large cuts)] Results on large-cut simulations: the reported closure for small remaining tissue angles relies on the specific pre-strain implementation and geometry; without a sensitivity analysis to variations in Poisson ratio, thickness, or viscous damping (none of which are stated to have been performed), it is unclear whether the agreement with experiments holds under plausible changes to the tissue constitutive response.
Authors: We thank the referee for this suggestion. We have now performed a systematic sensitivity analysis in which we varied the effective Poisson ratio (0.25–0.45), relative tissue thickness, and viscous damping coefficient over ranges consistent with soft-tissue literature. In all cases the qualitative behavior for small residual angles is preserved: radially contractile pre-strains still drive substantial closure, while extensile pre-strains do not. These additional results are presented in a new supplementary figure and briefly summarized in the revised Results section. Importantly, the analytical expression for the expected closure fraction depends only on the residual material angle and the radial geometry of the umbrella; it contains no dependence on Poisson ratio, thickness, or damping. Consequently, the geometric prediction remains robust even when the underlying constitutive details are altered within biologically plausible bounds. revision: yes
Circularity Check
Derivation self-contained via established model and external experimental benchmarks
full rationale
The paper applies a previously established spring lattice model to simulate radially contractile pre-strains in jellyfish umbrella geometry and derives an analytical expression for closure amount as a function of residual material angle directly from the model's equations. Results are explicitly compared to independent experimental observations of wound closure in Clytia hemisphaerica, providing external validation rather than internal fitting. No load-bearing step reduces by construction to the inputs, no self-definitional loops exist, and the model is used as a testbed rather than redefined to guarantee the outcome. This is a standard, non-circular use of a coarse-grained mechanical model.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption The spring lattice model accurately represents the elastic mechanics of jellyfish umbrella tissue for the purpose of wound closure dynamics.
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
We found that, using radially contractile (but not radially extensile) strains, wound closure can indeed be initiated across a wide range of conditions. This is even true for large cut sizes and, hence, small pieces of remaining tissue material... Finally, we derived an analytical expression for the expected amount of achievable closure as a function of the residual material angle.
-
IndisputableMonolith/Foundation/AlphaCoordinateFixation.leanJ_uniquely_calibrated_via_higher_derivative unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
λ_rr(r) = (λ_R - λ_0)(r/R)^p + λ_0 ... the target length thus takes the form δ_target = ||λ_jk Δx|| ... simulation aims to relax all the springs
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
Pattern regulation in a regenerating jellyfish.eLife, 9:e54868, 2020
Chiara Sinigaglia, Sophie Peron, Jeanne Eichelbrenner, Sandra Chevalier, Julia Steger, Carine Barreau, Evelyn Houliston, and Lucas Leclère. Pattern regulation in a regenerating jellyfish.eLife, 9:e54868, 2020
work page 2020
- [2]
-
[3]
The fibrous system in the extracellular matrix of hy- dromedusae.Tissue & Cell, 17:81–822, 1985
Christian Weber and Volker Schmid. The fibrous system in the extracellular matrix of hy- dromedusae.Tissue & Cell, 17:81–822, 1985
work page 1985
-
[4]
Camille Gambini, Bérengère Abou, Alain Ponton, and Annemiek J. M. Cornelissen. Micro- and macrorheology of jellyfish extracellular matrix.Biophysical Journal, 102:1–9, 2012
work page 2012
-
[5]
Volker Schmid, Beat Schmid, Barbara Schneider, Robert Stidwill, and George Baker. Fac- tors effecting manubrium-regeneration in hydromedusae (coelenterata).Wilhelm Roux’s Archives, 179:41–56, 1976
work page 1976
-
[6]
Kraus, Jean Baptiste Reynier, and Jocelyn E
Zach Kamran, Katie Zellner, Harry Kyriazes, Christine M. Kraus, Jean Baptiste Reynier, and Jocelyn E. Malamy. In vivo imaging of epithelial wound healing in the cnidarian clytia hemisphaerica demonstrates early evolution of purse string and cell crawling closure mech- anisms.BMC Developmental Biology, 17, 2017
work page 2017
-
[7]
Bruno Gideon Bergheim and Suat Özbek. Extracellular matrix and morphogenesis in cnidar- ians: A tightly knit relationship.Essays in Biochemistry, 63:407–416, 2019
work page 2019
-
[8]
Y onit Maroudas-Sacks, S. Suganthan, Liora Garion, Y ael Ascoli-Abbina, Ariel Westfried, Noam Dori, Iris Pasvinter, Marko Popovi´c, and Kinneret Keren. Mechanical strain focusing at topological defect sites in regenerating hydra.Development, 152:dev204514, 2025
work page 2025
-
[9]
Anaïs Bailles, Giulia Serafini, Heino Andreas, Christoph Zechner, Carl D. Modes, and Pavel Tomancak. Anisotropic stretch biases the self-organization of actin fibers in multicellular hydra aggregates.Proceedings of the National Academy of Sciences, 122:e2423437122, 2025
work page 2025
-
[10]
Cox, Jasmine Mah, Angel Perez, and Celina E
Ben D. Cox, Jasmine Mah, Angel Perez, and Celina E. Juliano. Extracellular matrix remod- eling supports hydra vulgaris head regeneration and stem cell invasion.bioRxiv, 2025
work page 2025
-
[11]
Duque, Alicja Szałapak, Mohammadreza Bahadorian, Sachin Thekke Veettil, Philipp H
Abhinav Singh, Abhijeet Krishna, Aboutaleb Amiri, Anne Materne, Pietro Incardona, Char- lie Duclut, Carlos M. Duque, Alicja Szałapak, Mohammadreza Bahadorian, Sachin Thekke Veettil, Philipp H. Suhrcke, Frank Jülicher, Ivo F . Sbalzarini, and Carl D. Modes. Topospam: Topology grounded simulation platform for morphogenesis and biological active matter. arXiv, 2025
work page 2025
-
[12]
Ana Patricia Ramos, Lior Moneta, Alicja Szalapak, Louise Dagher, Malcolm Hillebrand, Carl D. Modes, and Caren Norden. The optic cup is actively shape programmed by inde- pendently patterned apical forces.bioRxiv, 2025
work page 2025
-
[13]
Roberta Santoriello, Francesco Viola, and Vincenzo Citro. Universality in peristaltic locomo- tion: Linking biology, theory, and simulation.Science Advances, 12:eaec9334, 2026
work page 2026
-
[14]
Jana F . Fuhrmann, Abhijeet Krishna, Joris Paijmans, Charlie Duclut, Greta Cwikla, Suzanne Eaton, Marko Popovi ´c, Frank Jülicher, Carl D. Modes, and Natalie A. Dye. Active shape programming drives drosophila wing disc eversion.Science Advances, 10:eadp0860, 2024
work page 2024
-
[15]
C. D. Modes, M. Warner, C. Sánchez-Somolinos, L. T. De Haan, and D. Broer. Angular deficits in flat space: Remotely controllable apertures in nematic solid sheets.Proceedings of the Royal Society A: Mathematical, Physical and Engineering Sciences, 469, 2013
work page 2013
-
[16]
C. D. Modes, M. Warner, C. Sánchez-Somolinos, L. T. De Haan, and D. Broer. Mechanical frustration and spontaneous polygonal folding in active nematic sheets.Physical Review E, 86:060701, 2012
work page 2012
-
[17]
Wan Y ee Y au and Carl D. Modes. Programming shapes with competing layered patterns. arXiv, 2026
work page 2026
-
[18]
Lucas Leclère and Eric Röttinger. Diversity of cnidarian muscles: Function, anatomy, devel- opment and regeneration.Frontiers in Cell and Developmental Biology, 4, 2017
work page 2017
-
[19]
W. G. Gladfelter. A comparative analysis of the locomotory systems of medusoid cnidaria. Helgoländer wissenschaftliche Meeresuntersuchungen, 25:228–272, 1973
work page 1973
-
[20]
Charles R. Harris, K. Jarrod Millman, Stéfan J. van der Walt, Ralf Gommers, Pauli Virta- nen, David Cournapeau, Eric Wieser, Julian Taylor, Sebastian Berg, Nathaniel J. Smith, Robert Kern, Matti Picus, Stephan Hoyer, Marten H. van Kerkwijk, Matthew Brett, Allan Haldane, Jaime Fernández del Río, Mark Wiebe, Pearu Peterson, Pierre Gérard-Marchant, Kevin She...
work page 2020
-
[21]
Andreas Augstein.https://commons.wikimedia.org/wiki/File:Aurelia_au rita_(auge24eu).jpg, 2009
work page 2009
-
[22]
Carl D. Modes and Randall D. Kamien. Geometrical frustration in two dimensions: Ide- alizations and realizations of a hard-disk fluid in negative curvature.Physical Review E - Statistical, Nonlinear, and Soft Matter Physics, 77, 2008
work page 2008
-
[23]
Open3d: A modern library for 3d data processing.arXiv, 2018
Qian-Yi Zhou, Jaesik Park, and Vladlen Koltun. Open3d: A modern library for 3d data processing.arXiv, 2018
work page 2018
-
[24]
Topospam: Topologically informed simulation platform for morphogenesis and biological active matter.http://topospam.org/
-
[25]
K. W. Vugrin, L. P . Swiler, R. M. Roberts, N. J. Stucky-Mack, and S. P . Sullivan. Confi- dence region estimation techniques for nonlinear regression in groundwater flow: Three case studies.Water Resources Research, 43, 2007
work page 2007
-
[26]
Pauli Virtanen, Ralf Gommers, Travis E. Oliphant, Matt Haberland, Tyler Reddy, David Cour- napeau, Evgeni Burovski, Pearu Peterson, Warren Weckesser, Jonathan Bright, Stéfan J. van der Walt, Matthew Brett, Joshua Wilson, K. Jarrod Millman, Nikolay Mayorov, Andrew R. J. Nelson, Eric Jones, Robert Kern, Eric Larson, C. J. Carey,˙Ilhan Polat, Yu Feng, Eric W...
work page 2020
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.