Synchronization and metachronal waves in an array of eukaryotic flagella
Pith reviewed 2026-05-19 20:31 UTC · model grok-4.3
The pith
Larger arrays of flagella sustain stable metachronal waves with fixed nonzero phase shifts between neighbors.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
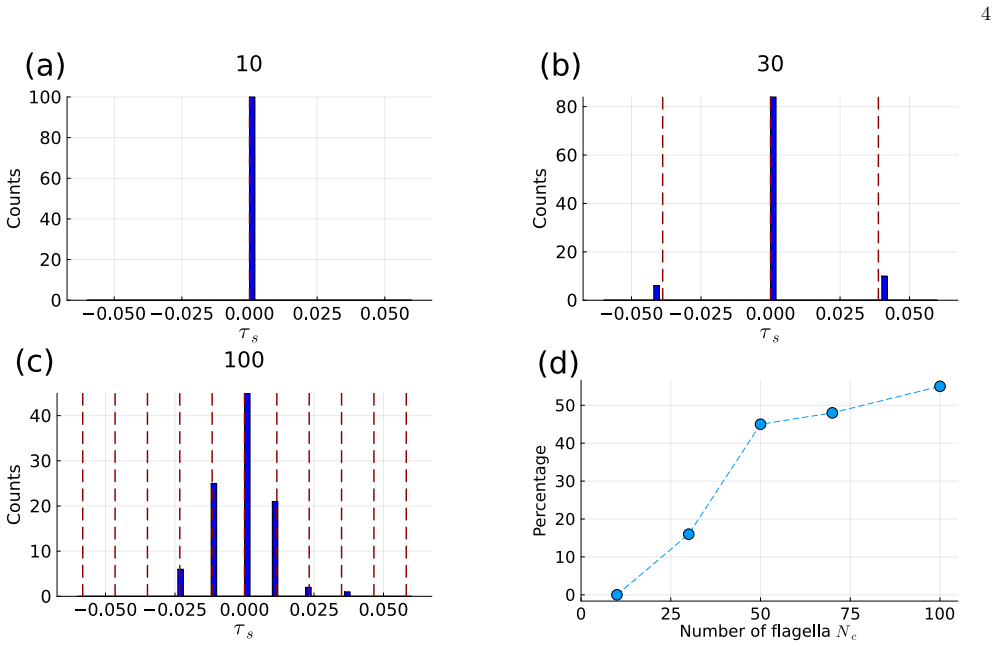
In contrast to a two-flagellum system where only in-phase synchronization is stable, larger arrays support stable metachronal waves with finite phase differences. Direct numerical simulations show that metachronal waves appear with increasing probability as the number of flagella increases. The analysis constructs a phase description for the array from the pair problem and shows that increasing system size enlarges the set of stable phase-locked modes under nearest-neighbor hydrodynamic coupling, thereby promoting metachronal-wave selection. A continuum description relates these collective states to advection and diffusion of the phase-difference field.
What carries the argument
The phase description for the array, constructed from the pair problem, which is used to analyze the stability of phase-locked states under nearest-neighbor hydrodynamic coupling.
If this is right
- Increasing the number of flagella enlarges the basin of attraction for metachronal waves relative to the in-phase state.
- The continuum limit maps the discrete phase-locked modes onto advection and diffusion of a phase-difference field.
- Direct simulations confirm that the probability of observing metachronal waves rises with array length.
- The framework explains how hydrodynamic coupling alone can generate robust traveling waves without additional biological regulation.
Where Pith is reading between the lines
- The same size-dependent enlargement of stable modes may operate in two-dimensional ciliary carpets, favoring coordinated waves for fluid transport.
- Varying the range of hydrodynamic coupling beyond nearest neighbors could shrink or expand the set of stable phase differences, offering a testable control parameter.
- If the phase model captures the essential mechanics, altering flagellar stiffness or spacing should shift the preferred phase lag in a predictable way.
Load-bearing premise
The phase dynamics of the entire array can be obtained by combining the pairwise interaction under the assumption of only nearest-neighbor hydrodynamic coupling.
What would settle it
A simulation or experiment in which metachronal waves with finite phase differences fail to appear at higher probability in larger arrays, or in which only the in-phase state remains stable regardless of array size, would disprove the claimed size-dependent selection.
Figures




read the original abstract
We investigate synchronization and metachronal-wave formation in a one-dimensional array of eukaryotic flagella using an elastohydrodynamic model. In contrast to a two-flagellum system, where only in-phase synchronization is stable, larger arrays are found to support stable metachronal waves with finite phase differences. Direct numerical simulations show that metachronal waves appear with increasing probability as the number of flagella increases. To explain this many-body effect, we construct a phase description for the array from that of the pair problem and analyze the stability of phase-locked states with nearest-neighbor hydrodynamic coupling. The analysis shows that increasing system size enlarges the set of stable phase-locked modes, thereby promoting metachronal-wave selection. A continuum description further relates these collective states to advection and diffusion of the phase-difference field. These results provide a simple theoretical framework for understanding how hydrodynamic interactions generate robust metachronal waves in flagellar arrays.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript studies synchronization and metachronal wave formation in a one-dimensional array of eukaryotic flagella governed by an elastohydrodynamic model. While a two-flagellum system supports only stable in-phase synchronization, direct numerical simulations of larger arrays show that metachronal waves with finite phase differences emerge with increasing probability as the number of flagella N grows. A phase-reduced description is constructed from the solved pair problem under nearest-neighbor hydrodynamic coupling; linear stability analysis of this model demonstrates that the number of stable phase-locked states (including those with nonzero constant phase difference) increases with system size. A continuum limit is derived that interprets the collective states in terms of advection and diffusion of the phase-difference field.
Significance. If the numerical observations and stability results hold, the work supplies a concrete many-body mechanism by which hydrodynamic interactions alone can select robust metachronal waves in flagellar arrays, with direct relevance to biological systems such as ciliated epithelia. The combination of full elastohydrodynamic DNS, phase reduction, and continuum analysis constitutes a clear strength; the explicit demonstration that the set of stable modes enlarges with N is a falsifiable prediction that can be tested in other models or experiments.
major comments (2)
- [§4] §4 (Direct numerical simulations): the reported increase in the probability of metachronal-wave states with N is central to the claim, yet the manuscript provides neither the full set of parameter values used in the elastohydrodynamic solver, nor discretization error estimates, nor a direct comparison of the two-flagellum limit against the independently solved pair problem. Without these, the robustness of the N-dependent trend cannot be assessed.
- [§5] §5 (Phase model and stability analysis): the array phase equations are obtained by truncating hydrodynamic interactions to nearest neighbors. Because the Stokes flow generated by each flagellum decays only as 1/r (or slower near a wall), next-nearest and longer-range torques remain non-negligible; their inclusion would modify the effective coupling function H(Δφ) and therefore the eigenvalues that determine stability of constant-phase-difference states. The conclusion that the number of stable modes grows with N may therefore be an artifact of the truncation rather than a generic many-body effect.
minor comments (2)
- [Figure 3] Figure 3: the color scale and axis labels for the phase-difference field in the continuum description should be stated explicitly in the caption so that the advection–diffusion interpretation can be read directly from the plot.
- [§5–§6] Notation: the symbol for the phase-difference field is introduced in the continuum section but used without redefinition in the stability analysis; a single consistent definition would improve readability.
Simulated Author's Rebuttal
We thank the referee for their constructive and detailed review, which has helped us improve the manuscript. We address each major comment below and have revised the paper accordingly to enhance the robustness and clarity of the presented results.
read point-by-point responses
-
Referee: [§4] §4 (Direct numerical simulations): the reported increase in the probability of metachronal-wave states with N is central to the claim, yet the manuscript provides neither the full set of parameter values used in the elastohydrodynamic solver, nor discretization error estimates, nor a direct comparison of the two-flagellum limit against the independently solved pair problem. Without these, the robustness of the N-dependent trend cannot be assessed.
Authors: We agree that these details are necessary to fully assess the robustness of the numerical observations. In the revised manuscript, we have added a dedicated section (or appendix) that lists all parameter values employed in the elastohydrodynamic solver. We have also included convergence studies with discretization error estimates obtained by systematically varying grid resolution and time-step size, confirming that the reported increase in metachronal-wave probability with N remains unchanged within the tested range. Finally, we now provide a direct side-by-side comparison between the two-flagellum synchronization behavior extracted from the array simulations and the independently solved pair problem, demonstrating quantitative agreement. These additions directly address the concern and strengthen the numerical evidence. revision: yes
-
Referee: [§5] §5 (Phase model and stability analysis): the array phase equations are obtained by truncating hydrodynamic interactions to nearest neighbors. Because the Stokes flow generated by each flagellum decays only as 1/r (or slower near a wall), next-nearest and longer-range torques remain non-negligible; their inclusion would modify the effective coupling function H(Δφ) and therefore the eigenvalues that determine stability of constant-phase-difference states. The conclusion that the number of stable modes grows with N may therefore be an artifact of the truncation rather than a generic many-body effect.
Authors: We appreciate the referee highlighting the slow decay of hydrodynamic interactions. The nearest-neighbor truncation is introduced to obtain a closed, analytically tractable phase model that extends the solved pair problem; this is a common modeling choice when the dominant coupling is local. In the revised manuscript we have added a paragraph discussing the relative magnitude of next-nearest-neighbor contributions, which are weaker by a factor of order 1/2 in the far-field Stokeslet approximation. We further show that the continuum limit of the phase-difference field remains qualitatively unchanged when a small longer-range term is included perturbatively. While a fully non-truncated many-body phase model would be computationally intensive, the existing analysis and the direct numerical simulations (which incorporate the full hydrodynamic interactions) both support that the enlargement of stable modes with N is a genuine many-body effect rather than solely an artifact of truncation. We have clarified these points and the limitations of the approximation in the text. revision: partial
Circularity Check
No circularity: derivation extends pair solution via explicit NN model without reduction to inputs
full rationale
The paper solves the two-flagellum elastohydrodynamic problem to obtain a phase description, then constructs the N-flagellum array model by imposing nearest-neighbor hydrodynamic coupling and performs linear stability analysis on the resulting phase equations. The reported growth in the number of stable phase-locked states (including finite-difference metachronal modes) with increasing N follows directly from the eigenvalues of the larger circulant coupling matrix; this is a mathematical property of the chain of oscillators and is not equivalent to any fitted quantity or self-referential definition. The NN truncation is an explicit modeling assumption stated in the text, not smuggled in via citation or forced by the target observation. No load-bearing step reduces the stability result to a tautology or to data fitted from the same metachronal-wave simulations.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Hydrodynamic interactions in the array can be approximated by nearest-neighbor coupling when constructing the phase description from the pair problem.
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
We construct a phase description for the array from that of the pair problem and analyze the stability of phase-locked states with nearest-neighbor hydrodynamic coupling. ... Substituting Γ(ψ) = −A sin(ψ + α) into Eq. (25) ... Λk = −2Aεω [cos ψ₀ cos α (1 − cos 2πk/Nc) − i sin ψ₀ sin α sin 2πk/Nc]
-
IndisputableMonolith/Foundation/DimensionForcing.leanreality_from_one_distinction unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
the phase difference satisfies ψ₀ = 2πn/Nc ... the stability condition becomes −Nc/4 < n < Nc/4
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
showsτ j(t)≈0, indicating in-phase synchronization. In Fig. 2(d), the steady state (t≳80) shows thatτ j(t) converges to a constant nonzero value, corresponding to a metachronal wave. In addition, stripe-like structures are observed in Fig. 2(d) during the intermediate stage (5≲t≲80). These stripes indicate that regions with relatively large phase differen...
-
[2]
S. L. Tamm, J. Cell Biol.55, 250 (1972)
work page 1972
-
[3]
D. R. Brumley, M. Polin, T. J. Pedley, and R. E. Gold- stein, Phys. Rev. Lett.109, 268102 (2012)
work page 2012
-
[4]
M. A. Sleigh, J. R. Blake, and N. Liron, Am. Rev. Respir. Dis.137, 726 (1988)
work page 1988
-
[5]
J. E. P. Lever, K. B. Turner, C. M. Fernandez, H. M. Leung, S. S. Hussain, R.-J. Shei, V. Y. Lin, S. E. Birket, K. K. Chu, G. J. Tearney, and G. M. Solomon, Am. J. Physiol.-Lung Cell. Mol. Physiol.327, L282 (2024)
work page 2024
- [6]
-
[7]
K. Y. Wan and R. N. Poon, Curr. Opin. Cell Biol.86, 102286 (2024)
work page 2024
-
[8]
C. Ringers, S. Bialonski, M. Ege, A. Solovev, J. N. Hansen, I. Jeong, B. M. Friedrich, and N. Jurisch-Yaksi, eLife12, e77701 (2023)
work page 2023
- [9]
-
[10]
N. Osterman and A. Vilfan, Proc. Natl. Acad. Sci. USA 108, 15727 (2011)
work page 2011
- [11]
-
[12]
G. J. Elfring and E. Lauga, Phys. Rev. Lett.103, 088101 (2009)
work page 2009
- [13]
- [14]
-
[15]
F. Meng, R. R. Bennett, N. Uchida, and R. Golestanian, Proc. Natl. Acad. Sci. USA118, e2102828118 (2021)
work page 2021
-
[16]
C. J. Brokaw, Biophys. J.12, 564 (1972)
work page 1972
- [17]
-
[18]
C. B. Lindemann, J. Theor. Biol.168, 175 (1994)
work page 1994
-
[19]
I. H. Riedel-Kruse, A. Hilfinger, J. Howard, and F. J¨ ulicher, HFSP J.1, 192 (2007)
work page 2007
-
[20]
P. V. Bayly and K. S. Wilson, Biophys. J.107, 1756 (2014)
work page 2014
-
[21]
B. Chakrabarti, S. F¨ urthauer, and M. J. Shelley, Proc. Natl. Acad. Sci. USA119, e2113539119 (2022)
work page 2022
-
[22]
R. E. Goldstein, E. Lauga, A. I. Pesci, and M. R. E. Proctor, Phys. Rev. Fluids1, 073201 (2016)
work page 2016
-
[23]
Y. Man, L. Koens, and E. Lauga, Europhys. Lett.116, 24002 (2016)
work page 2016
- [24]
-
[25]
N. Jung, W. K. Kim, and C. Hyeon, Phys. Rev. Research 7, 023029 (2025)
work page 2025
-
[26]
See Supplemental Material for time-evolution movies corresponding to the in-phase synchronized state, the metachronal-wave state, and the chevron pattern
-
[27]
D. A. Wiley, S. H. Strogatz, and M. Girvan, Chaos16, 015103 (2006)
work page 2006
-
[28]
P. Groisman, C. De Vita, J. Fern´ andez Bonder, and Y. Zhang, arXiv preprint arXiv:2506.03419 (2025)
- [29]
-
[30]
D. R. Brumley, N. Bruot, J. Kotar, R. E. Goldstein, P. Cicuta, and M. Polin, Phys. Rev. Fluids1, 081201 (2016)
work page 2016
- [31]
-
[32]
T. Wang, T. ul Islam, E. Steur, T. Homan, I. Aggarwal, P. R. Onck, J. M. J. den Toonder, and Y. Wang, Lab Chip24, 1573 (2024)
work page 2024
-
[33]
A. von Kenne, S. Schmelter, H. Stark, and M. B¨ ar, Phys. Rev. Research7, L012029 (2025)
work page 2025
-
[34]
Synchronization and metachronal waves in an array of eukaryotic flagella
Y. Wakahara and N. Uchida, Dataset and codes for “Synchronization and metachronal waves in an array of eukaryotic flagella”, https://doi.org/10.5281/zenodo.20209774
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.