Recognition: 2 theorem links
· Lean TheoremMechanical stress induced by the polymerisation of an active gel near a surface
Pith reviewed 2026-05-15 01:15 UTC · model grok-4.3
The pith
A hydrodynamic model of a compressible active gel shows that actin polymerization at the membrane can generate stresses that render a slightly corrugated membrane linearly unstable.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
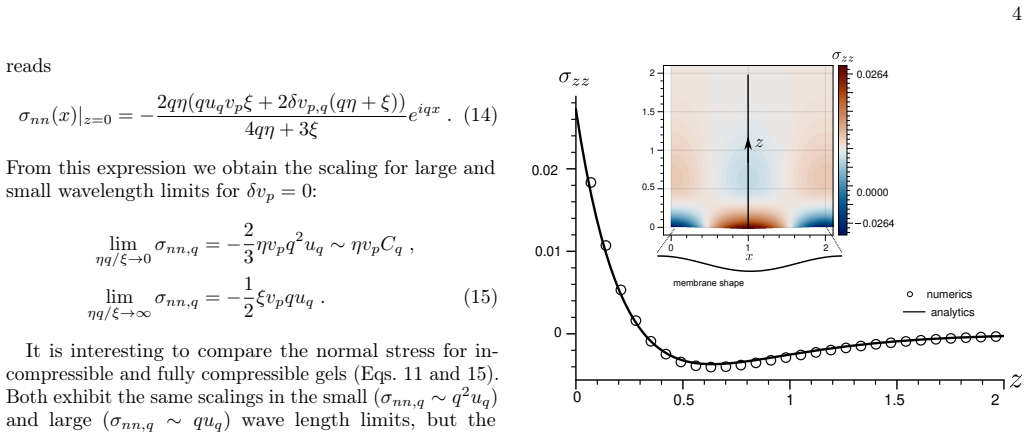
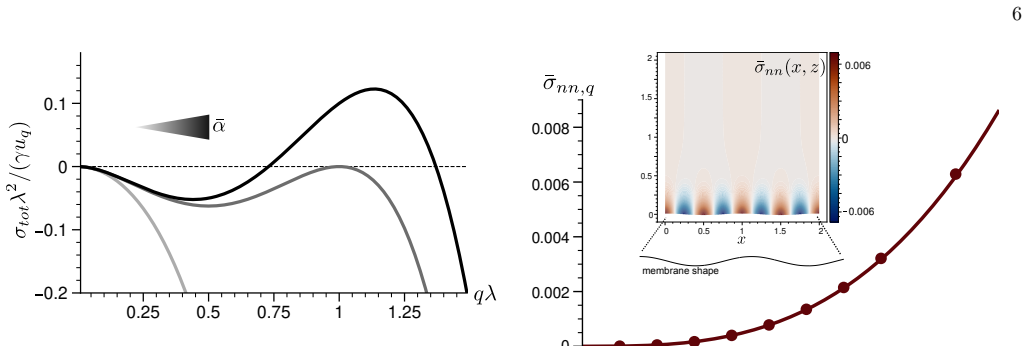
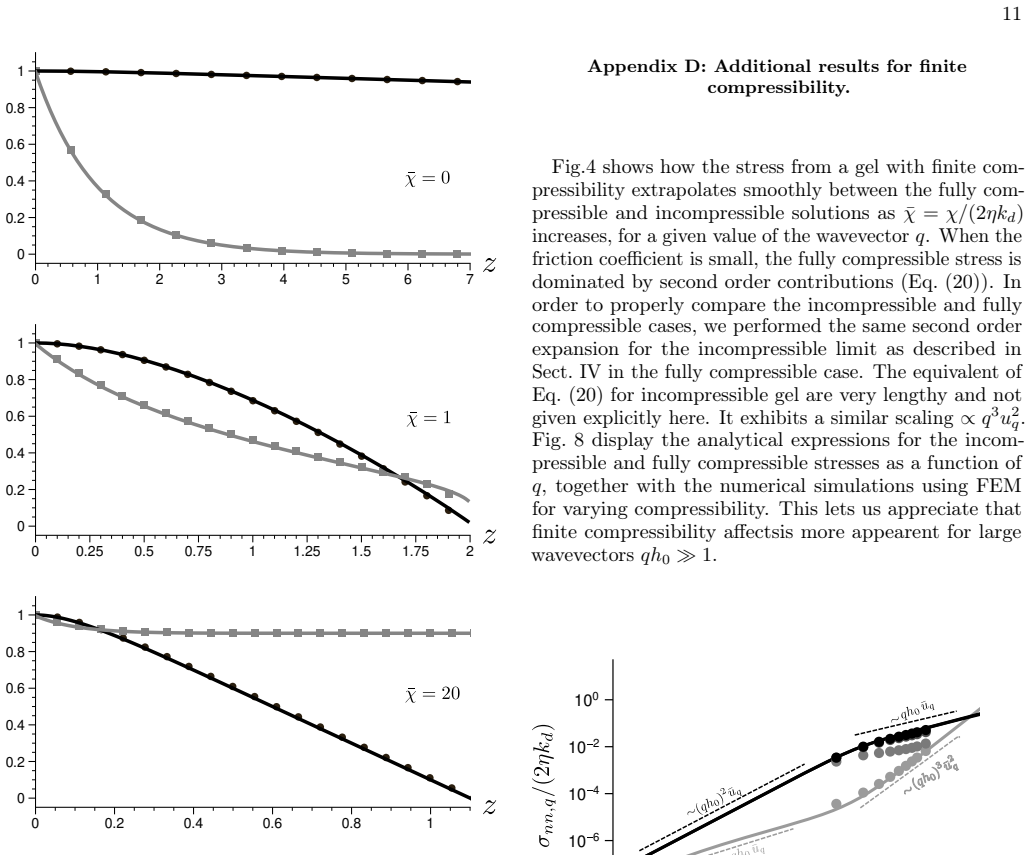
Analytical solutions in limiting regimes together with finite-element calculations in the general case provide a map of normal and tangential stresses on the membrane as functions of compressibility, interfacial friction and actin turnover, and they identify the conditions under which actin polymerization drives linear instability of the membrane.
What carries the argument
The hydrodynamic model of a compressible active gel that polymerizes at the membrane and turns over, evaluated at linear order in membrane corrugation amplitude.
If this is right
- Stresses increase with polymerization rate and decrease with higher compressibility or stronger friction.
- Linear instability occurs only inside a bounded window of turnover and friction values.
- Both normal and tangential force components act on the membrane and can cooperate to amplify corrugations.
- The same model can be continued into the weakly nonlinear regime to track saturation of the instability.
Where Pith is reading between the lines
- The instability threshold may set a natural length scale for cortical ripples or blebs in real cells.
- Adding membrane tension or curvature would shift the instability boundary and could stabilize or select particular wavelengths.
- The framework could be reused for other polymerizing active gels, such as those in bacterial cell walls or extracellular matrices.
Load-bearing premise
Membrane deformations remain small enough that a linear analysis in corrugation amplitude is valid and the gel obeys the stated hydrodynamic constitutive relations near the surface.
What would settle it
Direct measurement of whether small membrane corrugations grow or decay when actin turnover rate or membrane friction is varied across the predicted instability threshold.
Figures



read the original abstract
Actin flow in the cortical cytoskeleton underneath the cell membrane generates mechanical stresses that shape the cell surface. We study this mechanism using a hydrodynamic model of a compressible active gel polymerizing at the membrane and undergoing turnover. We determine how actin flow, density relaxation and friction of actin with the membrane generate stress on a corrugated membrane at the linear order in deformation. Analytical solutions in limiting regimes, combined with finite element methods in the general case, provide a map of normal and tangential stresses as functions of compressibility, interfacial friction and actin turnover, and determine the conditions under which actin polymerization can render the membrane linearly unstable. The non-linear regime is also briefly discussed.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript develops a hydrodynamic model of a compressible active gel that polymerizes at a membrane interface while undergoing turnover. It computes the normal and tangential stresses exerted on a corrugated membrane at linear order in the deformation amplitude, obtaining analytical expressions in limiting regimes of compressibility, friction, and turnover, and supplementing these with finite-element solutions in the general case. The central result is a parameter map that identifies the conditions under which actin polymerization renders the membrane linearly unstable.
Significance. If the central claims hold, the work supplies a concrete, parameter-dependent map of stresses and an explicit instability threshold for an active-gel-driven membrane corrugation. The combination of closed-form limits (which serve as internal benchmarks) and FEM numerics is a methodological strength that allows systematic exploration of the three free parameters (compressibility, interfacial friction coefficient, actin turnover rate). The results are directly relevant to cortical-actin-driven cell-surface patterning and rest on standard hydrodynamic constitutive relations without evident internal circularity.
major comments (2)
- [§3] §3 (linear stability analysis): the derivation of the instability threshold assumes that the membrane deformation remains small enough for linearization to be valid, yet no quantitative estimate is given for the critical corrugation amplitude at which nonlinear terms (e.g., curvature-dependent polymerization or geometric nonlinearities) would invalidate the threshold. This estimate is load-bearing for the claim that polymerization can render the membrane unstable.
- [§4.2] §4.2 (FEM implementation): the boundary conditions at the membrane-gel interface are stated to incorporate both normal and tangential stresses, but the precise discretization of the turnover term and the friction coefficient in the weak form is not shown; without this, it is difficult to verify that the numerical instability threshold converges to the analytical limits reported in §3.
minor comments (2)
- [Model section] The notation for the actin turnover rate (denoted variously as k or τ^{-1} in the text) should be unified and defined once in the model section.
- [Figure 5] Figure 5 caption: the color scale for the normal-stress map is not labeled with units; adding the appropriate stress units would improve readability.
Simulated Author's Rebuttal
We thank the referee for the positive assessment and constructive comments on our manuscript. We address each major comment below and will revise the manuscript accordingly.
read point-by-point responses
-
Referee: §3 (linear stability analysis): the derivation of the instability threshold assumes that the membrane deformation remains small enough for linearization to be valid, yet no quantitative estimate is given for the critical corrugation amplitude at which nonlinear terms (e.g., curvature-dependent polymerization or geometric nonlinearities) would invalidate the threshold. This estimate is load-bearing for the claim that polymerization can render the membrane unstable.
Authors: We agree that an estimate of the linear regime's validity range strengthens the central claim. In the revised manuscript we will add to §3 a quantitative estimate of the critical corrugation amplitude by comparing the magnitude of the leading nonlinear corrections (curvature-dependent polymerization and geometric nonlinearities in the stress balance) to the linear terms, using the same parameter values that define the instability threshold. This will be presented as an order-of-magnitude bound on the deformation amplitude below which the reported threshold remains valid. revision: yes
-
Referee: §4.2 (FEM implementation): the boundary conditions at the membrane-gel interface are stated to incorporate both normal and tangential stresses, but the precise discretization of the turnover term and the friction coefficient in the weak form is not shown; without this, it is difficult to verify that the numerical instability threshold converges to the analytical limits reported in §3.
Authors: We will include in the revised §4.2 the explicit weak-form expressions used in the finite-element implementation. The turnover term appears as a linear relaxation source in the continuity equation and is discretized with standard Galerkin weighting; the friction coefficient enters the tangential traction balance and is incorporated via a consistent penalty term on the interface. With these details added, the convergence of the numerical instability threshold to the analytical limits of §3 can be verified directly. revision: yes
Circularity Check
Derivation self-contained from hydrodynamic equations
full rationale
The paper derives normal and tangential stresses from the hydrodynamic equations of a compressible active gel with polymerization at the membrane and turnover, using standard constitutive relations. Analytical solutions are obtained in limiting regimes and finite-element methods are used in the general case to map stresses as functions of independent parameters (compressibility, interfacial friction, actin turnover). The linear instability condition follows directly from linear-order analysis in membrane corrugation amplitude. No step reduces by construction to a fitted quantity, self-citation chain, or ansatz smuggled via prior work; all inputs are external parameters and the outputs are explicit functions thereof, rendering the derivation self-contained against external benchmarks.
Axiom & Free-Parameter Ledger
free parameters (3)
- compressibility
- interfacial friction coefficient
- actin turnover rate
axioms (2)
- domain assumption The active gel can be described by a continuum hydrodynamic model with linear constitutive relations near the membrane.
- domain assumption Deformations are small enough for linearization in the corrugation amplitude.
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
We model the gel as a compressible Newtonian fluid satisfying the constitutive stress relation σ=(−P(ρ)+ηb∇·v)I+η(∇v+∇vT).
-
IndisputableMonolith/Foundation/RealityFromDistinction.leanreality_from_one_distinction unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
Analytical solutions in limiting regimes, combined with finite element methods in the general case, provide a map of normal and tangential stresses as functions of compressibility, interfacial friction and actin turnover.
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
G. Salbreux, G. Charras, and E. Paluch, Actin cortex mechanics and cellular morphogenesis, Trends in Cell Biology22, 536 (2012)
work page 2012
- [2]
-
[3]
H. Zhao, A. Pykäläinen, and P. Lappalainen, I-bar domain proteins: linking actin and plasma membrane dynamics, Current Opinion in Cell Biology23, 14 (2011)
work page 2011
-
[4]
N. Levernier and K. Kruse, Spontaneous formation of chaotic protrusions in a polymerizing active gel layer, New Journal of Physics22, 013003 (2020)
work page 2020
-
[5]
J. Xu, A. Palmer, and D. Wirtz, Rheology and microrhe- ology of semiflexible polymer solutions: Actin filament networks, Macromolecules31, 6486 (1998)
work page 1998
-
[7]
F. Jülicher, K. Kruse, J. Prost, and J.-F. Joanny, Active behavior of the cytoskeleton, Physics Reports449, 3 (2007)
work page 2007
- [8]
-
[9]
H.-Y. Lou, W. Zhao, X. Li, L. Duan, A. Powers, M. Aka- matsu, F. Santoro, A. F. McGuire, Y. Cui, D. G. Drubin, and B. Cui, Membrane curvature underlies actin reor- ganization in response to nanoscale surface topography, Proceedings of the National Academy of Sciences116, 23143 (2019)
work page 2019
-
[10]
F. Tsai, J. M. Henderson, Z. Jarin, E. Kremneva, Y. Senju, J. Pernier, O. Mikhajlov, J. Manzi, K. Ko- gan, C. Le Clainche, P. Lappalainen, and P. Bassereau, Activated I-BAR IRSp53 clustering controls the forma- tion of VASP-actin-based membrane protrusions, Science Advances8(2022)
work page 2022
-
[11]
W. Helfrich, Elastic properties of lipid bilayers: Theory and possible experiments, Zeitschrift für Naturforschung C28, 693 (1973)
work page 1973
-
[12]
M. Fritzsche, A. Lewalle, T. Duke, K. Kruse, and G. Char- ras, Analysis of turnover dynamics of the submembranous actin cortex, Molecular Biology of the Cell24, 757 (2013)
work page 2013
-
[13]
J. R. Kuhn and T. D. Pollard, Real-time measurements of actin filament polymerization by total internal reflection fluorescence microscopy, Biophysical Journal88, 1387 (2005). 8
work page 2005
-
[14]
T. D. Pollard and G. G. Borisy, Cellular motility driven by assembly and disassembly of actin filaments, Cell112, 453 (2003)
work page 2003
-
[15]
P. Bieling, H. T. Li, and M. P. Mullins, Force feedback controls motor activity and mechanical properties of self- assembling branched actin networks, Cell164, 115 (2016)
work page 2016
-
[16]
G. T. Charras, M. Coughlin, T. J. Mitchison, and L. Ma- hadevan, Life and times of a cellular bleb, Biophysical Journal94, 1836 (2008)
work page 2008
-
[17]
A. Cordes, H. Witt, A. Gallemí-Pérez, B. Brückner, F. Grimm, M. Vache, T. Oswald, J. Bodenschatz, D. Flor- mann, F. Lautenschläger, M. Tarantola, and A. Janshoff, Pre-stress of actin cortices is important for the viscoelastic response of living cells, bioRxiv 10.1101/783613 (2020), preprint, posted 2020-03-25
-
[18]
E. Fischer-Friedrich, Y. Toyoda, C. J. Cattin, D. J. Müller, A. A. Hyman, and F. Jülicher, Rheology of the active cell cortex in mitosis, Biophysical Journal111, 589 (2016)
work page 2016
-
[19]
A. Zgorski, R. W. Pastor, and E. Lyman, Surface shear viscosity and interleaflet friction from nonequilibrium simulations of lipid bilayers, Journal of Chemical Theory and Computation15, 6471 (2019)
work page 2019
-
[20]
J. E. Fitzgerald, R. M. Venable, R. W. Pastor, and L. E. R, Surface viscosities of lipid bilayers determined from equilibrium molecular dynamics simulations, Biophysical Journal122, 1094–1104 (2023)
work page 2023
-
[21]
M. P. Clausen, H. Colin-York, F. Schneider, C. Eggeling, and M. Fritzsche, Dissecting the actin cortex density and membrane-cortex distance in living cells by super- resolution microscopy, Journal of Physics D: Applied Physics50, 064002 (2017)
work page 2017
- [22]
-
[23]
A. Shamanskiy and B. Simeon, Mesh deformation tech- niques in fluid-structure interaction: robustness, accu- mulated distortion and computational efficiency, arXiv preprint arXiv:2006.14051 (2020)
-
[24]
J. Donea, A. Huerta, J.-P. Ponthot, and A. Rodríguez- Ferran, Arbitrary lagrangian–eulerian methods, inEn- cyclopedia of Computational Mechanics(John Wiley & Sons, Ltd, 2004) Chap. 14. 9 Appendix A: Contribution of bulk viscosity In the main text, we set bulk viscosityηb = 0and model the compressible actin gel with the constitutive maw σ = −P (ρ)I + η(∇v ...
work page 2004
-
[25]
Weak formulation To derive the weak formulation, the Stokes Eq.B1 is multiplied by a vector-valued test functionw∈ V V and the mass-conservation equation is multiplied by a scalar- valued test functionws ∈ V S, where VV and VS denote the spaces of vector- and scalar-valued test functions on Ω. After integration by parts, the momentum balance yields Fv = Z...
-
[26]
Free surface finite element procedure The problem is formulated as a free-boundary problem: the computational domain is not prescribed a priori but is determined self-consistently from the deformation of the free surface. The objective is to deform the domain iteratively through several steps (labeled byn) until the free boundary ∂Ωf coincides with the in...
-
[27]
Based on the chosen scales the diffu- sion constant is D = ¯Dh0vp
Effect of diffusion As mentioned before, diffusion is added to stabilize the numerical scheme by damping out arising short-length scale oscillations. Based on the chosen scales the diffu- sion constant is D = ¯Dh0vp. In order for diffusion to dampen potentially occurring numerical oscillations at the mesh length scalelmesh but not to influence our re- sul...
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.