Recognition: unknown
Prominent Signatures of Energy Transfer in Action-Detected Spectra of a Cyanobacterial Photosynthetic Protein
Pith reviewed 2026-05-07 14:43 UTC · model grok-4.3
The pith
Action-detected spectroscopy reveals energy transfer dynamics in a cyanobacterial protein because slow annihilation modifies the 1/N suppression.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
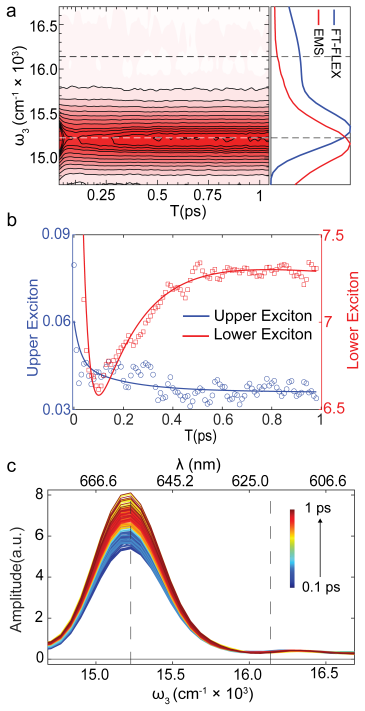
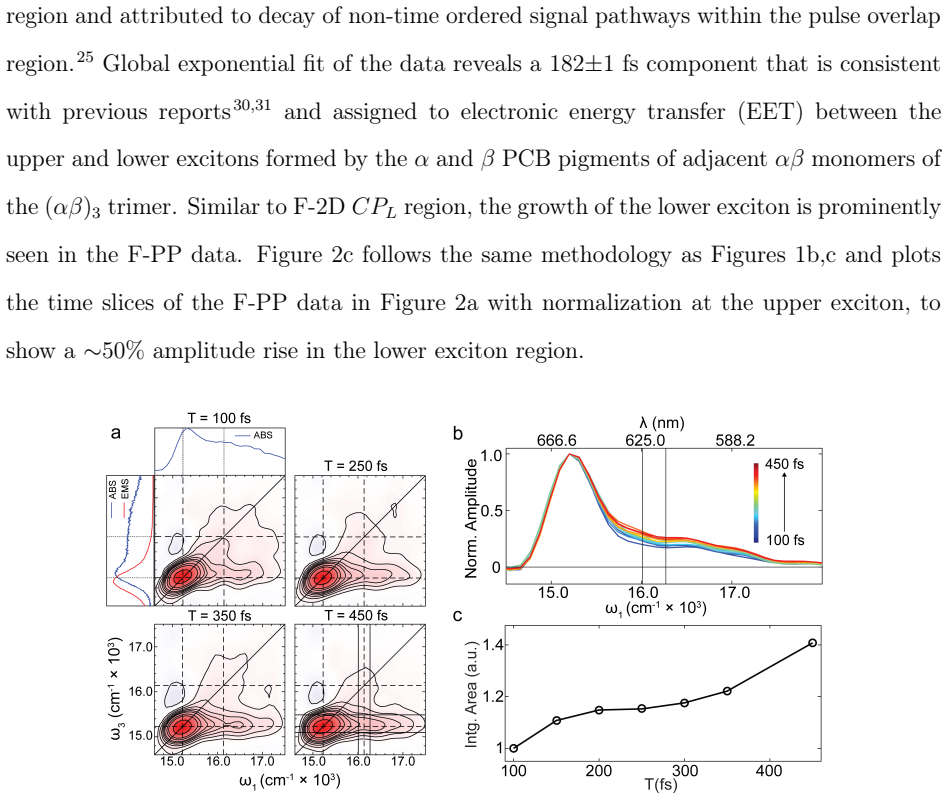
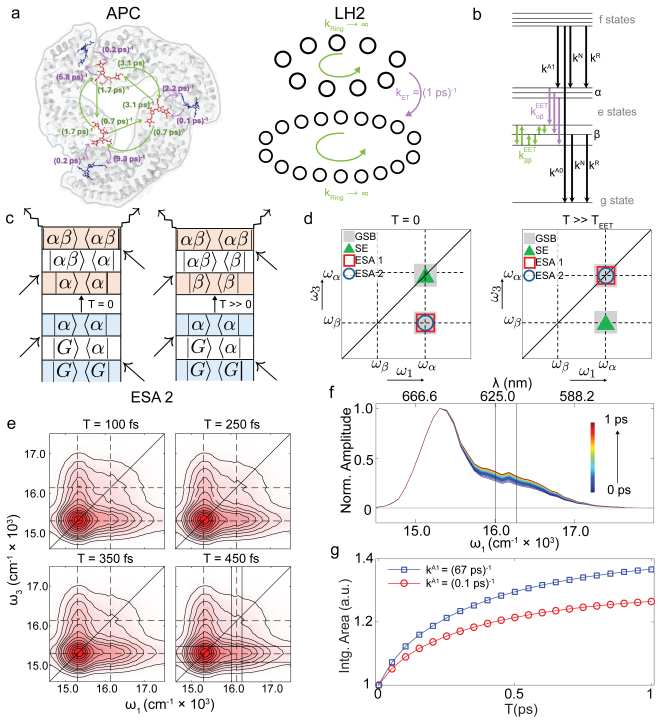
Measurements through both conventional and fluorescence 2DES show that the dynamics reported by F-2DES are quite prominent and comparable to conventional 2DES. Coarse-grained simulations combined with the experiments indicate that the 1/N limit argument, which assumes infinitely fast intra-exciton manifold equilibration, is modified in cyanobacterial proteins because of slow annihilation. The results suggest that action detection may in fact be well-suited to probe exciton diffusion across weakly coupled systems.
What carries the argument
The modification of the 1/N suppression in action-detected spectra arising from slow annihilation within the exciton manifold of the cyanobacterial protein.
Load-bearing premise
Intra-exciton manifold equilibration is infinitely fast, but this does not hold in cyanobacterial proteins due to slow annihilation.
What would settle it
A direct measurement of annihilation rates in the same cyanobacterial protein showing they are fast enough to restore the full 1/N suppression and eliminate the observed energy transfer signals.
Figures


read the original abstract
Action-detected two-dimensional electronic spectroscopy (A-2DES) could potentially be a versatile chemical tool with applicability across a range of photophysical observables such as photocurrent, photoionization, or fluorescence. However, a prominent absence of excited state energy/charge transfer dynamics signals in archetypal photosynthetic proteins has suggested severe limitations of A-2DES in probing large aggregates where sensitivity to excited state dynamics is proposed to go down as 1/N, where N is the aggregate size. We report measurements of energy transfer dynamics in a cyanobacterial protein through both conventional and fluorescence 2DES (F-2DES), where the dynamics reported by F-2DES is quite prominent and comparable to that measured by conventional 2DES. Analysis of our experiments combined with coarse-grained simulations of the spectra suggest that the 1/N limit argument, which assumes infinitely fast intra-exciton manifold equilibration, is modified in case of cyanobacterial proteins because of slow annihilation. Our results suggest that action detection may in fact be well-suited to probe exciton diffusion across weakly coupled systems.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript reports experimental measurements of energy transfer dynamics in a cyanobacterial photosynthetic protein using both conventional 2DES and fluorescence-detected action 2DES (F-2DES). It observes that the dynamics reported by F-2DES are prominent and comparable in strength to those from conventional 2DES, in contrast to the expected 1/N suppression for large aggregates. Coarse-grained simulations are used to interpret the results, suggesting that slow annihilation in these proteins modifies the assumption of infinitely fast intra-exciton manifold equilibration underlying the 1/N limit. The authors conclude that action detection may be well-suited to probe exciton diffusion across weakly coupled systems.
Significance. If the central claim holds, the work is significant because it provides direct experimental evidence challenging the presumed limitations of action-detected 2DES in photosynthetic aggregates and offers a mechanistic reinterpretation based on annihilation timescales. Credit is due for the side-by-side experimental comparison of conventional and action-detected spectra on the same sample and for employing simulations to connect observations to the modified 1/N picture. This could broaden the applicability of A-2DES to weakly coupled systems if the rate comparison is secured.
major comments (2)
- [Abstract and Simulations] The reinterpretation of the 1/N limit (abstract and simulation discussion) rests on the assertion that annihilation is slow compared to intra-manifold equilibration in cyanobacterial proteins. However, the manuscript provides no explicit quantitative check—such as a direct comparison of the chosen annihilation rate in the coarse-grained model to measured or literature intra-manifold relaxation rates for the protein—that confirms the slow-annihilation regime. Without this, the conventional 1/N suppression argument remains potentially intact, and the attribution of prominent F-2DES signals to the proposed mechanism is not fully secured. This is load-bearing for the central claim.
- [Methods and Results] §Methods and Results: full details on data analysis procedures, error estimation, baseline subtraction, and the specific numerical values for simulation parameters (e.g., annihilation rate, coupling strengths, relaxation timescales) are not provided. This prevents independent assessment of whether the observed comparability between F-2DES and conventional 2DES is robust or sensitive to analysis choices.
minor comments (2)
- [Figures] Figure captions and axis labels could more explicitly indicate which traces correspond to F-2DES versus conventional 2DES to aid immediate comparison.
- [Abstract] A brief statement of the specific cyanobacterial protein (e.g., its name or PDB code) in the abstract would improve clarity for readers unfamiliar with the system.
Simulated Author's Rebuttal
We thank the referee for their constructive and detailed comments, which have helped us strengthen the manuscript. We address each major point below and have made revisions where appropriate to improve clarity and support for our claims.
read point-by-point responses
-
Referee: [Abstract and Simulations] The reinterpretation of the 1/N limit (abstract and simulation discussion) rests on the assertion that annihilation is slow compared to intra-manifold equilibration in cyanobacterial proteins. However, the manuscript provides no explicit quantitative check—such as a direct comparison of the chosen annihilation rate in the coarse-grained model to measured or literature intra-manifold relaxation rates for the protein—that confirms the slow-annihilation regime. Without this, the conventional 1/N suppression argument remains potentially intact, and the attribution of prominent F-2DES signals to the proposed mechanism is not fully secured. This is load-bearing for the central claim.
Authors: We appreciate the referee highlighting this as a load-bearing aspect of the interpretation. The original manuscript selected the annihilation rate in the coarse-grained model from values reported in the literature for related photosynthetic aggregates, and the simulations demonstrate that this rate leads to the observed lack of 1/N suppression. However, we acknowledge that an explicit, side-by-side comparison to intra-manifold relaxation rates was not included. In the revised version, we have added a dedicated paragraph in the simulation discussion (and a supporting table in the SI) that compiles representative literature values for intra-exciton manifold equilibration times in cyanobacterial proteins and directly contrasts them with the annihilation timescale employed in our model. This addition confirms operation in the slow-annihilation regime relative to equilibration and thereby justifies the modification to the standard 1/N picture. We believe this addresses the concern without altering the central conclusions. revision: yes
-
Referee: [Methods and Results] §Methods and Results: full details on data analysis procedures, error estimation, baseline subtraction, and the specific numerical values for simulation parameters (e.g., annihilation rate, coupling strengths, relaxation timescales) are not provided. This prevents independent assessment of whether the observed comparability between F-2DES and conventional 2DES is robust or sensitive to analysis choices.
Authors: We agree that expanded methodological transparency is necessary for reproducibility. The revised manuscript now includes a substantially expanded Methods section that details the full data analysis pipeline, including the specific procedures used for baseline subtraction, error estimation (via replicate measurements and statistical analysis), and any peak-fitting or normalization steps. In addition, we have added a table in the Supplementary Information that lists all simulation parameters with their exact numerical values and literature sources. These changes allow independent evaluation of the robustness of the F-2DES versus conventional 2DES comparison. revision: yes
Circularity Check
No circularity; experimental data and interpretive modeling remain independent
full rationale
The paper reports new F-2DES measurements on a cyanobacterial protein that directly show prominent energy-transfer signals comparable to conventional 2DES. Coarse-grained simulations are invoked only to interpret why the conventional 1/N suppression is lifted (slow annihilation relative to intra-manifold relaxation). No equation, parameter fit, or claim reduces by construction to its own inputs; the central suggestion that action detection can probe exciton diffusion follows from the observed spectra rather than from a tautological redefinition or self-citation chain. Any self-citations present are peripheral and non-load-bearing.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption The proposed 1/N scaling of sensitivity to excited state dynamics in large aggregates for action-detected spectroscopy
Reference graph
Works this paper leans on
-
[1]
(2) Jonas, D. M. Vibrational and Nonadiabatic Coherence in 2D Electronic Spectroscopy, the Jahn–Teller Effect, and Energy Transfer.Annual Review of Physical Chemistry 2018,69, 327–352. (3) Policht, V. R.; Mittenzwey, H.; Dogadov, O.; Katzer, M.; Villa, A.; Li, Q.; Kaiser, B.; Ross, A. M.; Scotognella, F.; Zhu, X.; Knorr, A.; Selig, M.; Cerullo, G.; Dal Co...
2018
-
[2]
D.; Nuß, M.; Soavi, G.; Wang, G.; Ferrari, A
(11) Li, D.; Trovatello, C.; Conte, S. D.; Nuß, M.; Soavi, G.; Wang, G.; Ferrari, A. C.; Cerullo, G.; Brixner, T. Exciton–phonon coupling strength in single-layer MoSe2 at room temperature.Nature Communications2021,12. (12) Bruder, L.; Bangert, U.; Binz, M.; Uhl, D.; Vexiau, R.; Bouloufa-Maafa, N.; Dulieu, O.; Stienkemeier, F. Coherent multidimensional sp...
2012
-
[3]
A.; Konar, A.; Yu, Z.; Ptaszek, M.; Bocian, D
(18) Tiwari, V.; Matutes, Y. A.; Konar, A.; Yu, Z.; Ptaszek, M.; Bocian, D. F.; Holten, D.; Kirmaier, C.; Ogilvie, J. P. Strongly coupled bacteriochlorin dyad studied using phase- modulated fluorescence-detected two-dimensional electronic spectroscopy.Optics Ex- press2018,26, 22327. (19) Malý, P.; Lüttig, J.; Mueller, S.; Schreck, M. H.; Lambert, C.; Brix...
2002
-
[4]
J.; Edington, M
24 (32) Homoelle, B. J.; Edington, M. D.; Diffey, W. M.; Beck, W. F. Stimulated Photon-Echo and Transient-Grating Studies of Protein-Matrix Solvation Dynamics and Interexciton- StateRadiationlessDecayinαPhycocyaninandAllophycocyanin.The Journal of Phys- ical Chemistry B1998,102, 3044–3052. (33) Scholes, G. D.; Fleming, G. R. On the Mechanism of Light Harv...
2000
-
[5]
A.; Sauer, P
(45) Domínguez-Martín, M. A.; Sauer, P. V.; Kirst, H.; Sutter, M.; Bína, D.; Greber, B. J.; Nogales, E.; Polívka, T.; Kerfeld, C. A. Structures of a Phycobilisome in Light- Harvesting and Photoprotected States.Nature2022,609, 835–845. (46) Gonzalez-Migoni, S.; Jansen, T. L. C. Fluorescence-detected two-dimensional electronic 26 spectroscopy: A coarse-grai...
2026
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.