Chreode: A Cell World Model for One-Step Temporal Dynamics and Perturbation Prediction
Pith reviewed 2026-06-29 14:32 UTC · model grok-4.3
The pith
A one-step cell world model pretrained on developmental trajectories transfers to predict genetic perturbation responses.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
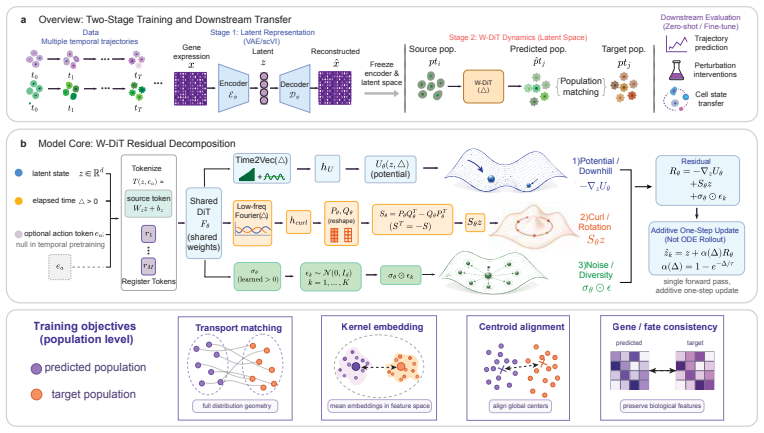
Chreode uses a structured residual transition operator to model action-conditioned cell-state transitions in one step, shifting distributional evolution to training time. Pretrained on 2.4M cells from mouse atlas, it serves as initialization for fine-tuning and as transferable embedding for perturbation models, showing that developmental dynamics aid CRISPR state shift prediction.
What carries the argument
The structured residual transition operator that decomposes cell-state evolution into downhill landscape flow, rotational in-tangent dynamics, and stochastic spread.
Load-bearing premise
The geometry and dynamics learned from normal developmental trajectories align sufficiently with the latent space of CRISPR perturbation responses to allow direct transfer without additional alignment or retraining.
What would settle it
Testing the pretrained model on a perturbation dataset whose cell states lie outside the developmental atlas trajectories yields no error reduction or worse performance than models trained from scratch on the perturbation data alone.
Figures

read the original abstract
Predicting how a cell will change its transcriptional state under a developmental signal or a genetic perturbation is the computational core of in-silico biology and the AI Virtual Cell program. Existing approaches either fit static control-to-treated maps that discard time, or solve multi-step ODE / Schr\"odinger-bridge problems on each dataset independently. We introduce Chreode, a one-step cell world model that predicts action-conditioned cell-state transitions through a structured residual transition operator. It shifts distributional evolution from inference time to training time, enabling single-pass generation while preserving a Waddington-inspired decomposition into downhill landscape flow, rotational in-tangent dynamics, and stochastic spread. The model is pretrained with a shared scVI encoder and a DiT-based dynamics backbone on a 2.4M-cell mouse embryonic atlas spanning 7 datasets. As a fine-tuning initialization, Chreode improves per-target Sinkhorn distance on Weinreb hematopoiesis and Veres islet differentiation over matched scratch models, PI-SDE, and PRESCIENT. As a transferable gene-state embedding for GEARS, the pretrained dynamics representation reduces shared-vocabulary DE20 mean squared error on Norman Perturb-seq from 0.2121 to 0.1858, a 12.4% relative improvement, without changing the GEARS training procedure. We interpret this transfer to perturbation prediction as evidence that pretrained developmental-trajectory dynamics encode differentiation primitives transferable to CRISPR-induced state shifts, since both involve cell-state transitions in a shared latent geometry. The pretrained backbone additionally produces zero-shot clonal fate scores on Weinreb that are competitive with strong dynamic-OT baselines.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper introduces Chreode, a one-step cell world model that uses a shared scVI encoder and DiT-based dynamics backbone pretrained on a 2.4M-cell mouse embryonic atlas. It claims this enables single-pass prediction of action-conditioned cell-state transitions via a structured residual transition operator (decomposing into downhill flow, rotational dynamics, and stochastic spread). When used as initialization, it improves Sinkhorn distances on Weinreb and Veres differentiation tasks over baselines; when used as a gene-state embedding inside unchanged GEARS, it reduces DE20 MSE on Norman Perturb-seq from 0.2121 to 0.1858 (12.4% relative gain). The authors interpret the transfer as evidence that developmental-trajectory dynamics encode transferable differentiation primitives applicable to CRISPR perturbations.
Significance. If the transfer result holds after proper controls, the work would demonstrate that large-scale pretraining on normal developmental trajectories can yield reusable dynamics representations for perturbation modeling, supporting the AI Virtual Cell agenda. The one-step formulation and Waddington-inspired decomposition are conceptually attractive, and the zero-shot clonal fate scores provide an additional falsifiable test. However, the current evidence does not yet isolate the contribution of the temporal dynamics component.
major comments (3)
- [Abstract; GEARS transfer experiment] Abstract and GEARS transfer results: the 0.2121→0.1858 DE20 MSE reduction is attributed to 'pretrained developmental-trajectory dynamics,' yet no ablation is reported that substitutes only the atlas-trained scVI encoder embeddings (without the residual transition operator or DiT pretraining) into the same GEARS pipeline. This control is required to establish that the gain arises from the learned dynamics rather than richer static gene representations from the 2.4M-cell corpus.
- [Abstract; interpretation paragraph] Abstract and results on transfer: the interpretation that the improvement demonstrates 'differentiation primitives transferable to CRISPR-induced state shifts' rests on the untested assumption that developmental and perturbation geometries overlap sufficiently for direct transfer. No alignment metric, latent-space visualization, or cross-domain distance analysis is provided to support this premise.
- [Abstract] Abstract: the reported numeric improvements (12.4% relative MSE reduction, competitive zero-shot clonal fate scores) are given without error bars, number of runs, or statistical significance tests, and full methods/data splits for the Norman Perturb-seq experiment are not detailed, preventing verification of the transfer claim.
minor comments (1)
- [Methods] The manuscript would benefit from explicit notation for the residual transition operator components (downhill, rotational, stochastic) and their relation to the DiT backbone in the methods section.
Simulated Author's Rebuttal
We thank the referee for the constructive comments. We address each major point below and will revise the manuscript to strengthen the claims with additional controls, analyses, and reporting details.
read point-by-point responses
-
Referee: [Abstract; GEARS transfer experiment] Abstract and GEARS transfer results: the 0.2121→0.1858 DE20 MSE reduction is attributed to 'pretrained developmental-trajectory dynamics,' yet no ablation is reported that substitutes only the atlas-trained scVI encoder embeddings (without the residual transition operator or DiT pretraining) into the same GEARS pipeline. This control is required to establish that the gain arises from the learned dynamics rather than richer static gene representations from the 2.4M-cell corpus.
Authors: We agree this ablation is required to isolate the dynamics contribution. We will add results using only the atlas-pretrained scVI encoder embeddings (no DiT or residual operator) in the GEARS pipeline and compare directly to the full Chreode initialization. revision: yes
-
Referee: [Abstract; interpretation paragraph] Abstract and results on transfer: the interpretation that the improvement demonstrates 'differentiation primitives transferable to CRISPR-induced state shifts' rests on the untested assumption that developmental and perturbation geometries overlap sufficiently for direct transfer. No alignment metric, latent-space visualization, or cross-domain distance analysis is provided to support this premise.
Authors: The performance transfer itself constitutes evidence of shared structure, but we acknowledge the need for explicit support. We will add UMAP visualizations of the joint latent space, compute cross-domain alignment metrics (e.g., average nearest-neighbor distances between developmental and perturbation trajectories), and include these in the revised results. revision: partial
-
Referee: [Abstract] Abstract: the reported numeric improvements (12.4% relative MSE reduction, competitive zero-shot clonal fate scores) are given without error bars, number of runs, or statistical significance tests, and full methods/data splits for the Norman Perturb-seq experiment are not detailed, preventing verification of the transfer claim.
Authors: We agree that statistical rigor and reproducibility details are essential. We will rerun the Norman experiments across multiple random seeds, report means with standard deviations and p-values, and expand the methods section with complete data splits, preprocessing, and training hyperparameters for the Perturb-seq transfer. revision: yes
Circularity Check
No circularity; derivation chain not reducible from provided text
full rationale
The abstract and reader's summary contain no equations, no self-citations, and no derivation steps that reduce a claimed prediction to its inputs by construction. The reported transfer gain (0.2121 to 0.1858 DE20 MSE) is presented as empirical evidence of transferable primitives, but this rests on an untested alignment assumption rather than a definitional or fitted-input loop. No load-bearing step matches any enumerated circularity pattern because no paper text is available to quote a reduction (e.g., no fitted parameter renamed as prediction or ansatz smuggled via citation). Per hard rules, absence of quotable reduction requires score 0 and empty steps list.
Axiom & Free-Parameter Ledger
Reference graph
Works this paper leans on
-
[1]
Generative Modeling via Drifting
Mingyang Deng, He Li, Tianhong Li, Yilun Du, and Kaiming He. Generative modeling via drifting.arXiv preprint arXiv:2602.04770,
work page internal anchor Pith review Pith/arXiv arXiv
-
[2]
David Ha and Jürgen Schmidhuber. World models.arXiv preprint arXiv:1803.10122,
work page internal anchor Pith review Pith/arXiv arXiv
-
[3]
Time2Vec: Learning a Vector Representation of Time
Seyed Mehran Kazemi, Rishab Goel, Sepehr Eghbali, Janahan Ramanan, Jaspreet Sahota, Sanjay Thakur, Stella Wu, Cathal Smyth, Pascal Poupart, and Marcus Brubaker. Time2vec: Learning a vector representation of time. InarXiv preprint arXiv:1907.05321,
work page internal anchor Pith review Pith/arXiv arXiv 1907
-
[4]
Dominik Klein, Jonas Simon Fleck, Daniil Bobrovskiy, Lea Zimmermann, Sören Becker, Alessandro Palma, Leander Dony, Alejandro Tejada-Lapuerta, Guillaume Huguet, Hsiu-Chuan Lin, Nadezhda Azbukina, Fátima Sanchís-Calleja, Theo Uscidda, Artur Szalata, Manuel Gander, Aviv Regev, Barbara Treutlein, J. Gray Camp, and Fabian J. Theis. CellFlow enables generative ...
-
[5]
Branched Schrödinger bridge matching
Sophia Tang, Yinuo Zhang, Alexander Tong, and Pranam Chatterjee. Branched Schrödinger bridge matching. arXiv preprint arXiv:2506.09007,
-
[6]
Christina V
Verified via paper-search-mcp Google Scholar (2026-05-07); Semantic Scholar does not index this arXiv preprint at lookup time, so verify-bib returns a false-positive title mismatch. Christina V . Theodoris, Ling Xiao, Anant Chopra, Mark D. Chaffin, Zeina R. Al Sayed, Matthew C. Hill, Helene Mantineo, Elizabeth M. Brydon, Zexian Zeng, X. Shirley Liu, and P...
2026
-
[7]
Improving and generalizing flow-based generative models with minibatch optimal transport
Alexander Tong, Kilian Fatras, Nikolay Malkin, Guillaume Huguet, Yanlei Zhang, Jarrid Rector-Brooks, Guy Wolf, and Yoshua Bengio. Improving and generalizing flow-based generative models with minibatch optimal transport.arXiv preprint arXiv:2302.00482,
work page internal anchor Pith review Pith/arXiv arXiv
-
[8]
doi: 10.1038/s42256-025-01150-3. 11 C. H. Waddington.The Strategy of the Genes. George Allen & Unwin,
-
[9]
Jin Wang, Kun Zhang, Li Xu, and Erkang Wang
Xaira Therapeutics, bioRxiv 2026.03.18.712807. Jin Wang, Kun Zhang, Li Xu, and Erkang Wang. Quantifying the Waddington landscape and biological paths for development and differentiation.Proceedings of the National Academy of Sciences,
2026
-
[10]
[2021]; used for both time-transition and fate evaluation Veres islet as released with Tang et al
data release mouse development atlas Weinreb hematopoiesis GEO GSE140802 subset as released with Yeo et al. [2021]; used for both time-transition and fate evaluation Veres islet as released with Tang et al
2021
-
[11]
7-timepoint differentiation Norman Perturb-seq GEO GSE133344 [Norman et al., 2019] CellStream EMT, MOSTA shipped with Ling et al
2019
-
[12]
The scVI framework [Lopez et al., 2018] is used under the scvi-tools BSD-3 license
appendix velocity-consistency evaluation Baseline software: PRESCIENT [Yeo et al., 2021] under its GitHub license; BranchSBM [Tang et al., 2025] under its GitHub license; CellFlow [Klein et al., 2025a] under its release license; CellOT 14 [Bunne et al., 2023] and scGen [Lotfollahi et al., 2019] under their respective open-source licenses. The scVI framewo...
2021
-
[13]
“NFE” counts the number of network forward passes per (source cell, ∆, action) query. Wall-clock and FLOPs are reported as median over 60 Weinreb d2→d6 queries with 8 warmup queries discarded; FLOPs from torch.profiler. Method NFE / query ms / query GFLOPs / query PRESCIENT∼88 194.33 0.07 CellFlow 50–100 ODE410.36— Chreode 164.79 0.90 Chreode resolves a q...
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.