Spatio-Temporal Dynamics of Nucleo-Cytoplasmic Transport
Pith reviewed 2026-05-24 02:21 UTC · model grok-4.3
The pith
Spatial buildup of RanGEF near the nuclear envelope raises nuclear Ran levels by its own transport rules.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
The model yields steady-state profiles, relaxation times, and nuclear-to-cytoplasmic ratios that depend on spatial dynamics and heterogeneity. Specifically, the spatial nonuniformity of RanGEF, particularly its proximity to the nuclear envelope, increases the Ran content in the nucleus. RanGEF's accumulation near the nuclear envelope arises from its intrinsic dynamics as a nuclear cargo transported by the Ran cycle itself.
What carries the argument
The Ran cycle, in which RanGEF promotes GTP exchange on Ran inside the nucleus while spatial transport moves components across the nuclear envelope.
If this is right
- Nuclear-to-cytoplasmic molecule ratios depend on the spatial distribution of RanGEF.
- Relaxation times of the transport system vary with molecular spatial heterogeneity inside the nucleus.
- Steady-state profiles of Ran cycle components are shaped by proximity to the nuclear envelope.
- RanGEF positioning is self-organized by the same cycle it regulates.
Where Pith is reading between the lines
- Disrupting the envelope-proximal accumulation of RanGEF could reduce transport efficiency in conditions that alter nuclear geometry.
- Similar spatial self-organization may operate in other GTPase-regulated systems where cargo positioning feeds back on activity.
- Live-cell imaging of RanGEF gradients under controlled nuclear shapes could directly test the predicted accumulation mechanism.
Load-bearing premise
The model uses chosen reaction rates, diffusion coefficients, and a fixed nuclear geometry that produce the observed RanGEF spatial profiles as an outcome of the transport rules rather than from separate measurements.
What would settle it
An experiment that measures RanGEF distribution in live cells and finds no enrichment near the nuclear envelope, or that relocates RanGEF away from the envelope and observes no drop in nuclear Ran levels.
Figures





read the original abstract
Nucleocytoplasmic transport is essential for cellular function, presenting a canonical example of rapid molecular sorting inside cells. It consists of a coordinated interplay between import/export of molecules in/out the cell nucleus. Here, we investigate the role of spatio-temporal dynamics of the nucleocytoplasmic transport and its regulation. We develop a biophysical model that captures the main features of the nucleocytoplasmic transport, in particular, its regulation through the Ran cycle. Our model yields steady-state profiles for the molecular components of the Ran cycle, their relaxation times, as well as the nuclear-to-cytoplasmic molecule ratio. We show that these quantities are affected by their spatial dynamics and heterogeneity within the nucleus. Specifically, we find that the spatial nonuniformity of Ran Guanine Exchange Factor (RanGEF) - particularly its proximity to the nuclear envelope - increases the Ran content in the nucleus. We further show that RanGEF's accumulation near the nuclear envelope results from its intrinsic dynamics as a nuclear cargo, transported by the Ran cycle itself. Overall, our work highlights the critical role of molecular spatial dynamics in cellular processes, and proposes new avenues for theoretical and experimental inquiries into the nucleocytoplasmic transport.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript develops a biophysical model of nucleocytoplasmic transport that incorporates the Ran GTPase cycle to generate steady-state spatial profiles of its molecular components, relaxation times to steady state, and nuclear-to-cytoplasmic concentration ratios. It concludes that spatial nonuniformity of RanGEF, specifically its accumulation near the nuclear envelope, elevates nuclear Ran levels and that this accumulation itself emerges from RanGEF's transport as a nuclear cargo within the same cycle.
Significance. If the reported effect of RanGEF localization on nuclear Ran content proves robust, the work would illustrate how spatial heterogeneity can modulate transport efficiency and could motivate targeted experiments on RanGEF positioning. The absence of experimental validation or parameter-sensitivity tests, however, currently limits the strength of this contribution.
major comments (2)
- [Abstract] Abstract: the central claim that RanGEF nuclear-envelope accumulation 'results from its intrinsic dynamics as a nuclear cargo, transported by the Ran cycle itself' is presented without demonstration that the outcome is independent of the specific (unspecified) reaction rates, diffusion coefficients, and fixed nuclear geometry; the reported nuclear Ran increase may therefore be a direct consequence of those modeling choices rather than an emergent prediction.
- [Model formulation] Model formulation (inferred from abstract and results): the steady-state profiles and N/C ratios are outputs of a system whose spatial nonuniformity of RanGEF is not constrained by independent measurements; without a sensitivity analysis or comparison to measured nuclear-to-cytoplasmic ratios, the load-bearing assertion that proximity to the envelope increases nuclear Ran rests on untested parameter values.
minor comments (1)
- [Abstract] Abstract: the phrase 'yields steady-state profiles... as well as the nuclear-to-cytoplasmic molecule ratio' would benefit from explicit statement of which molecular species are included in the ratio.
Simulated Author's Rebuttal
We thank the referee for the constructive comments. We agree that parameter sensitivity and robustness to modeling choices require explicit demonstration and will add these analyses in revision. Below we respond point-by-point to the major comments.
read point-by-point responses
-
Referee: [Abstract] Abstract: the central claim that RanGEF nuclear-envelope accumulation 'results from its intrinsic dynamics as a nuclear cargo, transported by the Ran cycle itself' is presented without demonstration that the outcome is independent of the specific (unspecified) reaction rates, diffusion coefficients, and fixed nuclear geometry; the reported nuclear Ran increase may therefore be a direct consequence of those modeling choices rather than an emergent prediction.
Authors: We accept that the abstract and main text do not yet demonstrate independence from specific parameter values or geometry. The accumulation arises self-consistently from the coupled transport and reaction terms without being imposed as a boundary condition, but to establish robustness we will add a dedicated sensitivity section (varying diffusion coefficients, rate constants, and nuclear radius within biologically plausible ranges) and report the conditions under which the envelope accumulation and elevated nuclear Ran persist. The fixed spherical geometry is a standard modeling choice; we will note its limitations and test modest shape perturbations. revision: yes
-
Referee: [Model formulation] Model formulation (inferred from abstract and results): the steady-state profiles and N/C ratios are outputs of a system whose spatial nonuniformity of RanGEF is not constrained by independent measurements; without a sensitivity analysis or comparison to measured nuclear-to-cytoplasmic ratios, the load-bearing assertion that proximity to the envelope increases nuclear Ran rests on untested parameter values.
Authors: We agree that the spatial nonuniformity of RanGEF emerges from the model dynamics rather than being fitted to data, and that direct comparison to measured N/C ratios is needed. In revision we will (i) perform and present a systematic parameter-sensitivity analysis, (ii) tabulate predicted N/C ratios against published experimental values for Ran and related components, and (iii) clarify which parameters are taken from literature versus chosen for numerical stability. These additions will make the dependence on parameter choices transparent. revision: yes
- Direct experimental validation of the predicted RanGEF localization effect would require new live-cell imaging or perturbation experiments that lie outside the scope of the present theoretical study.
Circularity Check
No circularity: results are direct numerical outputs of a standard reaction-diffusion model with chosen parameters and geometry.
full rationale
The paper constructs a biophysical model of the Ran cycle with explicit reaction rates, diffusion coefficients, and fixed nuclear geometry, then numerically computes steady-state profiles and ratios. The reported RanGEF accumulation near the envelope and consequent nuclear Ran increase are outputs of those equations and boundary conditions, not reductions of the target quantities to themselves by definition or self-citation. No load-bearing self-citation, uniqueness theorem, or ansatz smuggling is described. This is the normal case of a simulation study whose predictions depend on its inputs; the derivation chain does not collapse by construction.
Axiom & Free-Parameter Ledger
free parameters (1)
- reaction rates and diffusion coefficients
axioms (1)
- domain assumption The Ran cycle consists of standard GTPase reactions with importin/exportin mediation
Reference graph
Works this paper leans on
-
[1]
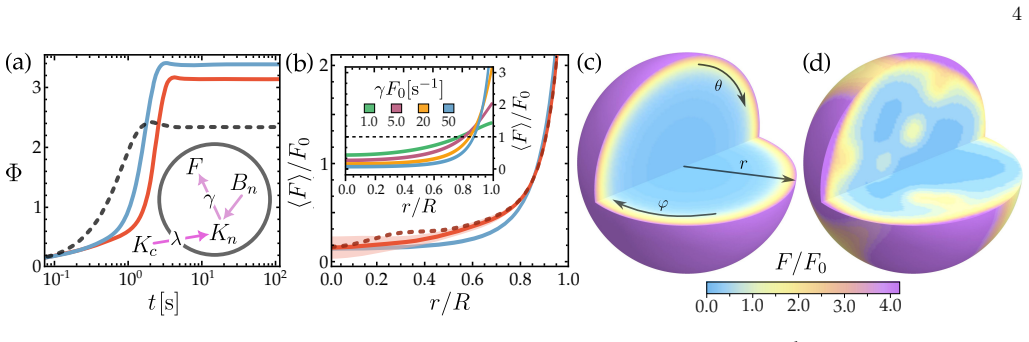
which is located primarily at the nuclear boundary. A reduction in Φ has been experimentally observed by decreasing either the levels of heterochromatin markers or HP1 [37–40]. Furthermore, disrupting nuclear lamin, as seen in progerin patient cell lines where both lamin disruption and heterochromatin alterations are evident, also reduces Φ [37–40], being...
-
[2]
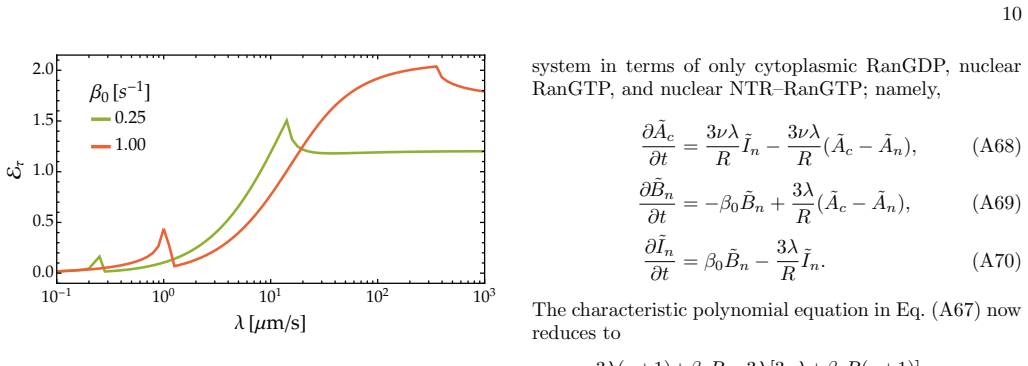
Radial solutions in Laplace space Provided that the initial conditions for the nuclear con- centrations are uniform, their steady-state solutions will remain spherically symmetric as the cytoplasmic concen- trations are treated as spatially homogeneous. The nu- clear concentration of RanGDP can be described by T ∂An ∂t = 1 r2 ∂ ∂r r2 ∂An ∂r − kαAn(r, t), ...
-
[3]
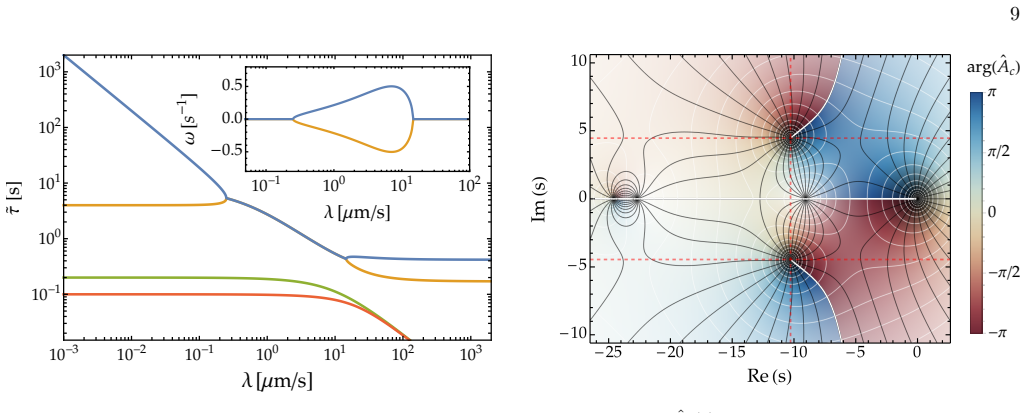
Steady-state concentration profiles The steady-state concentration profiles can be derived by final value theorem for the Laplace transform, which tells us that if the steady-state value exists, then lim t→∞ f(t) = lim s→0 s ˆF (s) where ˆF (s) is the Laplace transform of the function f(t), provided that all poles of s ˆF (s) are strictly stable or lie in...
-
[4]
Nuclear-to-cytoplasmic ratio of molecules The number of cytoplasmic molecules in the form of RanGDP and NTR–RanGTP at time t are given by cA(t) = Vc Ac(t), and cI(t) = Vc Ic(t), (A41) respectively, whereas the net number of nuclear RanGDP, nA(t), RanGTP, nB(t), and NTR–RanGTP, nI(t) are given by the integral of their corresponding concentration fields ove...
-
[5]
Relaxation times to steady-state Before computing the corresponding relaxation times of the system to steady-state from the Laplace space so- lutions in Eqs. (A13), (A14), (A18), (A22) and (A25), it is instructive to calculate the relaxation times for the fast diffusion case (d → ∞), which leads to rapid homogeniza- tion of nuclear concentrations. In this...
-
[6]
Steady-state profiles — the shell case Using Eq. (A1), the steady-state equation for nuclear RanGDP can be written as follows: 1 r2 ∂ ∂r h r2 ∂ ¯Aso n ∂r i = 0, 0 < r ≤ 1−δ, 1 r2 ∂ ∂r h r2 ∂ ¯Asi n ∂r i = ks α ¯Asi n, 1−δ < r ≤ 1. (B5) where ks α = R2α0/[d(1 − δ3)], while ¯Asi n and ¯Aso n denote the concentrations inside and outside the shell, re...
-
[7]
Steady-state profiles — the ball case From Eq. (A1), the steady-state equation for nuclear RanGDP is given by 1 r2 ∂ ∂r h r2 ∂ ¯Abo n ∂r i = 0, ε < r ≤ 1, 1 r2 ∂ ∂r h r2 ∂ ¯Abi n ∂r i = kb α ¯Abi n , 0 < r ≤ ε. (B23) where kb α = R2α0/(dε3), whilst ¯Abi n and ¯Abo n are the con- centrations inside and outside the spherical ball, respec- tively. Th...
-
[8]
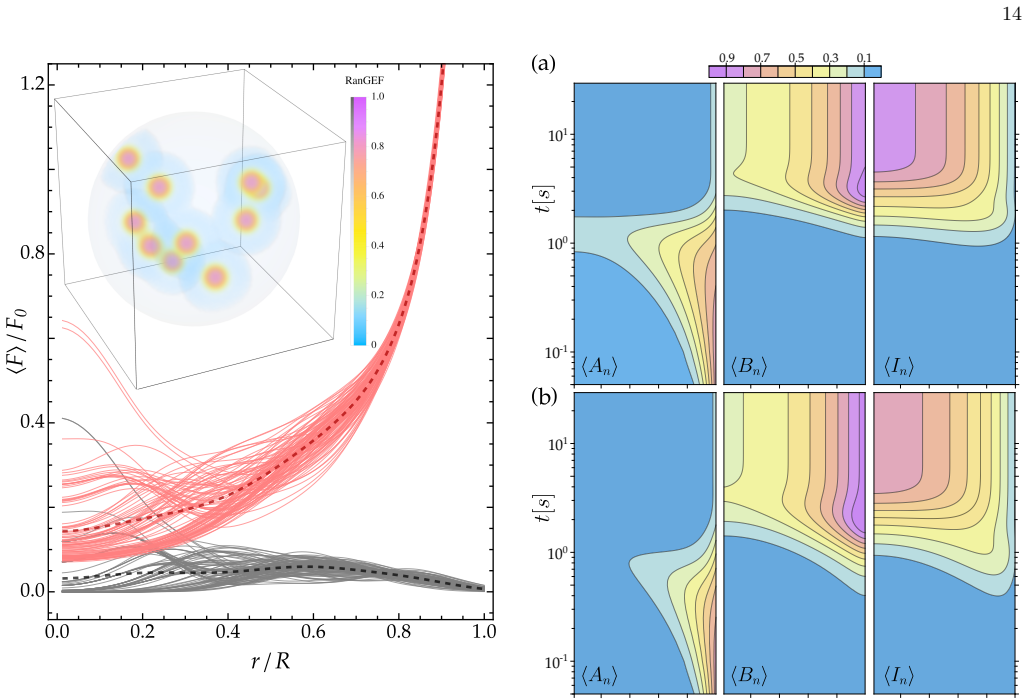
Net number of molecules and their ratios By integrating the concentration fields over the nu- clear volume, we can compute the respective net number of molecules. Using Eq. (A42), total number of nuclear 101 100 10-1 101 100 10-1 0.2 00.4 0.6 0.8 0.2 0.4 0.6 0.8 0.2 0.4 0.6 0.8 1 0.9 0.7 0.5 0.3 0.1 FIG. 11. Angular average of the nuclear RanGDP ⟨An⟩, Ran...
-
[9]
B. Alberts, A. Johnson, J. Lewis, M. Raff, K. Roberts, and P. Walter, Molecular Biology of the Cell , 5th ed. (Garland Science, 2008)
work page 2008
-
[10]
S. R. Wente and M. P. Rout, The nuclear pore complex and nuclear transport, Cold Spring Harbor Perspectives in Biology 2, a000562 (2010)
work page 2010
-
[11]
B. L. Timney, B. Raveh, R. Mironska, J. M. Trivedi, S. J. Kim, D. Russel, S. R. Wente, A. Sali, and M. P. Rout, Simple rules for passive diffusion through the nuclear pore complex, Journal of Cell Biology 215, 57 (2016)
work page 2016
-
[12]
O. Keminer and R. Peters, Permeability of single nuclear pores, Biophysical Journal 77, 217 (1999)
work page 1999
-
[13]
D. Mohr, S. Frey, T. Fischer, T. G¨ uttler, and D. G¨ orlich, Characterisation of the passive permeability barrier of nuclear pore complexes, The EMBO Journal 28, 2541 (2009)
work page 2009
-
[14]
E. E. Benarroch, Nucleocytoplasmic transport, Neurol- ogy 92, 757 (2019)
work page 2019
-
[15]
I. G. Macara, Transport into and out of the nucleus, Microbiology and Molecular Biology Reviews 65, 570 (2001)
work page 2001
-
[16]
B. Cautain, R. Hill, N. de Pedro, and W. Link, Compo- nents and regulation of nuclear transport processes, The FEBS Journal 282, 445 (2015)
work page 2015
-
[17]
T. Jovanovic-Talisman and A. Zilman, Protein transport by the nuclear pore complex: Simple biophysics of a com- plex biomachine, Biophysical Journal 113, 6 (2017)
work page 2017
-
[18]
T. Zheng and A. Zilman, Self-regulation of the nuclear pore complex enables clogging-free crowded transport, Proceedings of the National Academy of Sciences 120, 10.1073/pnas.2212874120 (2023). 16
-
[19]
W. Yang, J. Gelles, and S. M. Musser, Imaging of single- molecule translocation through nuclear pore complexes, Proceedings of the National Academy of Sciences 101, 12887 (2004)
work page 2004
-
[20]
K. Ribbeck, U. Kutay, E. Paraskeva, and D. G¨ orlich, The translocation of transportin–cargo complexes through nuclear pores is independent of both ran and energy, Cur- rent Biology 9, 47 (1999)
work page 1999
-
[21]
D. G¨ orlich and U. Kutay, Transport between the cell nu- cleus and the cytoplasm, Annual Review of Cell and De- velopmental Biology 15, 607 (1999)
work page 1999
-
[22]
T. Cavazza and I. Vernos, The rangtp pathway: From nucleo-cytoplasmic transport to spindle assembly and be- yond, Frontiers in Cell and Developmental Biology 3, 10.3389/fcell.2015.00082 (2016)
-
[23]
U. Kubitscheck, D. Grunwald, A. Hoekstra, D. Rohleder, T. Kues, J. P. Siebrasse, and R. Peters, Nuclear transport of single molecules, The Journal of Cell Biology 168, 233 (2005)
work page 2005
-
[24]
B. W. Hoogenboom, L. E. Hough, E. A. Lemke, R. Y. Lim, P. R. Onck, and A. Zilman, Physics of the nu- clear pore complex: Theory, modeling and experiment, Physics Reports 921, 1 (2021)
work page 2021
-
[25]
Joseph, Ran at a glance, Journal of Cell Science 119, 3481 (2006)
J. Joseph, Ran at a glance, Journal of Cell Science 119, 3481 (2006)
work page 2006
-
[26]
H. Y. Li, D. Wirtz, and Y. Zheng, A mechanism of cou- pling rcc1 mobility to rangtp production on the chro- matin in vivo, Journal of Cell Biology 160, 635 (2003)
work page 2003
-
[27]
J. R. Hutchins, W. J. Moore, F. E. Hood, J. S. Wilson, P. D. Andrews, J. R. Swedlow, and P. R. Clarke, Phos- phorylation regulates the dynamic interaction of rcc1 with chromosomes during mitosis, Current Biology 14, 1099 (2004)
work page 2004
-
[28]
I. Cushman, D. Stenoien, and M. S. Moore, The dynamic association of rcc1 with chromatin is modulated by ran- dependent nuclear transport, Molecular Biology of the Cell 15, 245 (2004)
work page 2004
-
[29]
E. Hitakomate, F. E. Hood, H. S. Sanderson, and P. R. Clarke, The methylated n-terminal tail of rcc1 is required for stabilisation of its interaction with chromatin by ran in live cells, BMC Cell Biology 11, 43 (2010)
work page 2010
-
[30]
M. J. Seewald, C. K¨ orner, A. Wittinghofer, and I. R. Vet- ter, Rangap mediates gtp hydrolysis without an arginine finger, Nature 415, 662 (2002)
work page 2002
-
[31]
For transport proteins like Importin- β, which utilize an adaptor protein, Importin- α, to attach to the cargo, the energy expenditure involves two molecules of GTP
-
[32]
M. V. Nachury and K. Weis, The direction of transport through the nuclear pore can be inverted, Proceedings of the National Academy of Sciences 96, 9622 (1999)
work page 1999
-
[33]
Thermodynamic paradigm for solution demixing inspired by nuclear transport in living cells, Physical Review Let- ters 118, 158101 (2017)
work page 2017
-
[34]
D. G¨ orlich, M. J. Seewald, and K. Ribbeck, Character- ization of ran-driven cargo transport and the rangtpase system by kinetic measurements and computer simula- tion, The EMBO Journal 22, 1088 (2003)
work page 2003
-
[35]
A. E. Smith, Systems analysis of ran transport, Science 295, 488 (2002)
work page 2002
-
[36]
R. B. Kopito and M. Elbaum, Reversibility in nucleocyto- plasmic transport, Proceedings of the National Academy of Sciences 104, 12743 (2007)
work page 2007
-
[37]
R. B. Kopito and M. Elbaum, Nucleocytoplasmic trans- port: A thermodynamic mechanism, HFSP Journal 3, 130 (2009)
work page 2009
-
[38]
B. L. Timney, J. Tetenbaum-Novatt, D. S. Agate, R. Williams, W. Zhang, B. T. Chait, and M. P. Rout, Simple kinetic relationships and nonspecific competition govern nuclear import rates in vivo, The Journal of Cell Biology 175, 579 (2006)
work page 2006
-
[39]
K. Ribbeck and D. G¨ orlich, Kinetic analysis of transloca- tion through nuclear pore complexes, The EMBO Journal 20, 1320 (2001)
work page 2001
-
[40]
G. Riddick and I. G. Macara, The adapter importin- α provides flexible control of nuclear import at the expense of efficiency, Molecular Systems Biology 3, 10.1038/msb4100160 (2007)
- [41]
- [42]
-
[43]
J. Wu, A. H. Corbett, and K. M. Berland, The intracellu- lar mobility of nuclear import receptors and nls cargoes, Biophysical Journal 96, 3840 (2009)
work page 2009
-
[44]
A. Abu-Arish, P. Kalab, J. Ng-Kamstra, K. Weis, and C. Fradin, Spatial distribution and mobility of the ran gtpase in live interphase cells, Biophysical Journal 97, 2164 (2009)
work page 2009
-
[45]
J. B. Kelley, S. Datta, C. J. Snow, M. Chatterjee, L. Ni, A. Spencer, C.-S. Yang, C. Cube˜ nas-Potts, M. J. Matu- nis, and B. M. Paschal, The defective nuclear lamina in hutchinson-gilford progeria syndrome disrupts the nucle- ocytoplasmic ran gradient and inhibits nuclear localiza- tion of ubc9, Molecular and Cellular Biology 31, 3378 (2011)
work page 2011
- [46]
-
[47]
M. Chatterjee and B. M. Paschal, Disruption of the ran system by cysteine oxidation of the nucleotide ex- change factor rcc1, Molecular and Cellular Biology 35, 566 (2015)
work page 2015
-
[48]
N. Dworak, D. Makosa, M. Chatterjee, K. Jividen, C. Yang, C. Snow, W. C. Simke, I. G. Johnson, J. B. Kel- ley, and B. M. Paschal, A nuclear lamina-chromatin-ran gtpase axis modulates nuclear import and dna damage signaling, Aging Cell 18, 10.1111/acel.12851 (2019)
-
[49]
J. M. Casolari, C. R. Brown, S. Komili, J. West, H. Hi- eronymus, and P. A. Silver, Genome-wide localization of the nuclear transport machinery couples transcriptional status and nuclear organization, Cell 117, 427 (2004)
work page 2004
-
[50]
A. Bizhanova and P. D. Kaufman, Close to the edge: Het- erochromatin at the nucleolar and nuclear peripheries, Biochimica et Biophysica Acta (BBA) - Gene Regulatory Mechanisms 1864, 194666 (2021)
work page 2021
-
[51]
A. Zidovska, The self-stirred genome: large-scale chro- matin dynamics, its biophysical origins and implications, Current Opinion in Genetics and Development 61, 83 (2020)
work page 2020
-
[52]
A. R. Lowe, J. H. Tang, J. Yassif, M. Graf, W. Y. Huang, J. T. Groves, K. Weis, and J. T. Liphardt, Importin- β modulates the permeability of the nuclear pore complex in a ran-dependent manner, eLife 4, 10.7554/eLife.04052 (2015)
-
[53]
M. Winey, D. Yarar, T. H. Giddings, and D. N. Mas- tronarde, Nuclear pore complex number and distribu- 17 tion throughout the saccharomyces cerevisiae cell cycle by three-dimensional reconstruction from electron micro- graphs of nuclear envelopes, Molecular Biology of the Cell 8, 2119 (1997)
work page 1997
-
[54]
S. S. Ashwin, T. Nozaki, K. Maeshima, and M. Sa- sai, Organization of fast and slow chromatin revealed by single-nucleosome dynamics, Proceedings of the National Academy of Sciences 116, 19939 (2019)
work page 2019
-
[55]
J. Min´ e-Hattab and I. Chiolo, Complex chromatin motions for dna repair, Frontiers in Genetics 11, 10.3389/fgene.2020.00800 (2020)
-
[56]
G. M. Oliveira, A. Oravecz, D. Kobi, M. Maroquenne, K. Bystricky, T. Sexton, and N. Molina, Precise measure- ments of chromatin diffusion dynamics by modeling us- ing gaussian processes, Nature Communications 12, 6184 (2021)
work page 2021
-
[57]
R. Oshidari, K. Mekhail, and A. Seeber, Mobility and repair of damaged dna: Random or directed?, Trends in Cell Biology 30, 144 (2020)
work page 2020
-
[58]
R. Barth, K. Bystricky, and H. A. Shaban, Coupling chro- matin structure and dynamics by live super-resolution imaging, Science Advances 6, 10.1126/sciadv.aaz2196 (2020)
-
[59]
A. Zidovska, D. A. Weitz, and T. J. Mitchison, Micron- scale coherence in interphase chromatin dynamics, Pro- ceedings of the National Academy of Sciences 110, 15555 (2013)
work page 2013
-
[60]
D. Saintillan, M. J. Shelley, and A. Zidovska, Exten- sile motor activity drives coherent motions in a model of interphase chromatin, Proceedings of the National Academy of Sciences 115, 11442 (2018)
work page 2018
-
[61]
A. Mahajan, W. Yan, A. Zidovska, D. Saintillan, and M. J. Shelley, Euchromatin activity enhances segregation and compaction of heterochromatin in the cell nucleus, Physical Review X 12, 041033 (2022)
work page 2022
-
[62]
Here we do not ex- plicitly consider the interactions with NTF2
RanGDP is transported into the nucleus by binding to Nuclear Transport Factor 2 (NTF2). Here we do not ex- plicitly consider the interactions with NTF2. We assume that the rate of nucleotide exchange for Ran is not lim- ited by NTF2, assuming the RanGDP molecule dissoci- ates rapidly from that complex once it enters the nucleus; though its dissociation me...
-
[63]
T. Tachibana, M. Hieda, Y. Miyamoto, S. Kose, N. Imamoto, and Y. Yoneda, Recycling of importin alpha from the nucleus is suppressed by loss of rcc1 function in living mammalian cells., Cell Structure and Function 25, 115 (2000)
work page 2000
-
[64]
B. Banerjee, D. Bhattacharya, and G. Shivashankar, Chromatin structure exhibits spatio-temporal hetero- geneity within the cell nucleus, Biophysical Journal 91, 2297 (2006)
work page 2006
-
[65]
R. S. Sankhala, R. K. Lokareddy, S. Begum, R. A. Pum- roy, R. E. Gillilan, and G. Cingolani, Three-dimensional context rather than nls amino acid sequence determines importin α subtype specificity for rcc1, Nature Commu- nications 8, 979 (2017)
work page 2017
-
[66]
K. J. Burns, G. M. Vasil, J. S. Oishi, D. Lecoanet, and B. P. Brown, Dedalus: A flexible framework for numer- ical simulations with spectral methods, Physical Review Research 2, 023068 (2020)
work page 2020
-
[67]
T. Tachibana, N. Imamoto, H. Seino, T. Nishimoto, and Y. Yoneda, Loss of rcc1 leads to suppression of nu- clear protein import in living cells., Journal of Biological Chemistry 269, 24542 (1994)
work page 1994
-
[68]
F. E. Hood and P. R. Clarke, Rcc1 isoforms differ in their affinity for chromatin, molecular interactions and regulation by phosphorylation, Journal of Cell Science 120, 3436 (2007)
work page 2007
- [69]
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.