Robustness of Boolean networks to update modes: an application to hereditary angioedema
Pith reviewed 2026-05-22 13:26 UTC · model grok-4.3
The pith
The hereditary angioedema interaction graph contains subgraphs that activate in an alternating block-parallel mode with a constantly updated core and two alternating subsets, exhibiting robustness or instability relative to classical update
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
Subgraphs within the hereditary angioedema gene interaction graph activate through an alternating block-parallel mode featuring one core that is constantly updated and two complementary subsets that alternate their updating, and these subgraphs display structural robustness or instability when compared to classical periodic update modes.
What carries the argument
The alternating block-parallel update mode with one core constantly updated and two complementary subsets alternating, applied to subgraphs of the hereditary angioedema interaction graph.
If this is right
- The identified subgraphs maintain consistent activation under the alternating block-parallel schedule.
- Robust subgraphs indicate network features that remain stable across changes in update timing.
- Unstable subgraphs mark locations where small shifts in update order can disrupt coordinated gene activity.
- Direct comparison with classical periodic modes isolates which structural traits are independent of the update schedule.
Where Pith is reading between the lines
- Similar alternating-update subgraphs may exist in other familial-disease networks and could be searched for by the same structural criteria.
- Varying the assignment of genes to core versus alternating groups in simulation would test how sensitive the robustness result is to the initial partitioning.
- If the pattern generalizes, it supplies a concrete way to rank candidate intervention points by their update-mode stability.
Load-bearing premise
The particular interaction graph chosen and the division of genes into the constantly updated core and the two alternating subsets correctly reflect the actual regulatory logic in hereditary angioedema.
What would settle it
A simulation of the described subgraphs under the alternating block-parallel mode that produces activation patterns inconsistent with observed disease biology or that loses its reported robustness after modest rewiring of the graph edges would falsify the central claim.
Figures


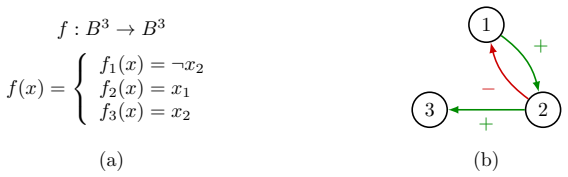







read the original abstract
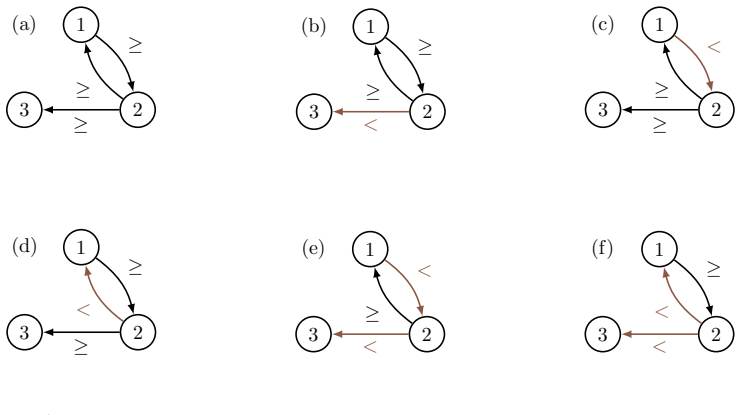
Many familial diseases are caused by genetic accidents, which affect both the genome and its epigenetic environment, expressed as an interaction graph between the genes as that involved in one familial disease we shall study, the hereditary angioedema. The update of the gene states at the vertices of this graph (1 if a gene is activated, 0 if it is inhibited) can be done in multiple ways, well studied over the last two decades: Parallel, sequential, block-sequential, block-parallel, random, etc. We will study a particular graph, related to the familial disease proposed as an example, which has subgraphs which activate in an intricate manner (\emph{i.e.}, in an alternating block-parallel mode, with one core constantly updated and two complementary subsets of genes alternating their updating), of which we will study the structural aspects, robust or unstable, in relation to some classical periodic update modes.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript studies the robustness of Boolean networks to update modes by analyzing a specific interaction graph associated with hereditary angioedema. It identifies subgraphs that activate in an alternating block-parallel mode (one core constantly updated, two complementary subsets alternating) and compares their structural robustness or instability to classical periodic modes such as parallel and sequential updates.
Significance. If the central claims hold, the work would contribute to the discrete-mathematics literature on Boolean-network dynamics by providing a concrete biological application that links update-mode sensitivity to disease-related regulatory graphs. The focus on structural properties rather than simulation alone offers a potentially falsifiable framework for assessing robustness across update schedules.
major comments (2)
- [Abstract and §1] Abstract and §1: the central claim that the hereditary-angioedema graph contains subgraphs exhibiting structural robustness (or instability) under the described alternating block-parallel mode rests on an unvalidated choice of graph and an unmotivated partitioning of vertices into a constant core plus two alternating blocks; no derivation from primary biological data or cross-validation against known regulatory interactions is supplied, so the disease-specific interpretation does not follow from the mathematical properties alone.
- [§3] §3 (or the section presenting the update-mode definitions): the alternating block-parallel schedule is introduced as 'intricate' without an explicit formal definition (e.g., no equation specifying the periodic schedule, the size of the core, or the complementarity condition on the alternating subsets), preventing verification that the reported robustness properties are independent of the particular partitioning chosen.
minor comments (2)
- [Figures] Figure captions and notation: ensure that every gene label appearing in the interaction graph is defined in a table or legend so that the core-versus-alternating assignment can be inspected without external lookup.
- [References] References: add citations to prior work on block-parallel update modes in Boolean networks (e.g., the original definitions of block-sequential and block-parallel schedules) to situate the alternating variant.
Simulated Author's Rebuttal
We thank the referee for the constructive comments. We address each major point below and indicate the revisions made to strengthen the manuscript.
read point-by-point responses
-
Referee: [Abstract and §1] Abstract and §1: the central claim that the hereditary-angioedema graph contains subgraphs exhibiting structural robustness (or instability) under the described alternating block-parallel mode rests on an unvalidated choice of graph and an unmotivated partitioning of vertices into a constant core plus two alternating blocks; no derivation from primary biological data or cross-validation against known regulatory interactions is supplied, so the disease-specific interpretation does not follow from the mathematical properties alone.
Authors: The hereditary angioedema interaction graph is presented as a concrete illustrative example drawn from the literature on the disease to motivate the mathematical study of update-mode robustness, rather than as a model derived directly from primary experimental data within this work. The partitioning into a constant core and two complementary alternating blocks is chosen based on the observed structure of the graph that supports the alternating block-parallel dynamics. We have revised the abstract and §1 to clarify the illustrative nature of the biological application and to emphasize that the robustness results are structural and mathematical. We agree that additional cross-validation against primary data would strengthen the disease link but consider it beyond the discrete-mathematics scope of the paper. revision: partial
-
Referee: [§3] §3 (or the section presenting the update-mode definitions): the alternating block-parallel schedule is introduced as 'intricate' without an explicit formal definition (e.g., no equation specifying the periodic schedule, the size of the core, or the complementarity condition on the alternating subsets), preventing verification that the reported robustness properties are independent of the particular partitioning chosen.
Authors: We agree that an explicit formal definition is required for clarity and reproducibility. In the revised manuscript we have added a precise definition in §3, including a mathematical equation for the periodic schedule, the fixed core vertex set, and the two complementary alternating subsets together with their update rules. This formalization makes the robustness properties verifiable independently of any specific partitioning. revision: yes
Circularity Check
No significant circularity; analysis applies standard update-mode methods to a chosen input graph
full rationale
The paper takes as given a particular interaction graph related to hereditary angioedema and analyzes the activation patterns and robustness of its subgraphs under alternating block-parallel and classical periodic update modes. No equations or claims reduce by construction to fitted parameters, self-definitions, or load-bearing self-citations; the graph and its partitioning are explicit modeling inputs, and the reported structural properties follow from direct examination of that graph rather than from any circular renaming or imported uniqueness theorem. The derivation chain is therefore self-contained against external Boolean-network benchmarks.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption The gene interaction graph for hereditary angioedema is known and fixed.
- ad hoc to paper Alternating block-parallel update with a constant core and two complementary alternating subsets is a relevant mode for this network.
Lean theorems connected to this paper
-
IndisputableMonolith/Foundation/ArrowOfTime.leanarrow_from_z unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
robustness to variations in synchronism... if changes are observed in terms of gene expression... unlikely to stem from the influence of biological clocks
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
J. Aracena, J. Demongeot, ´E. Fanchon, and M. Montalva. On the number of different dy- namics in Boolean networks with deterministic update schedules.Mathematical Biosciences, 242:188–194, 2013
work page 2013
-
[2]
J. Aracena, J. Demongeot, ´E. Fanchon, and M. Montalva. On the number of update digraphs and its relation with the feedback arc sets and tournaments.Discrete Applied Mathematics, 161:1345–1355, 2013
work page 2013
-
[3]
J. Aracena, J. Demongeot, and E. Goles. Positive and negative circuits in discrete neural networks.IEEE Transactions on Neural Networks, 15:77–83, 2004
work page 2004
-
[4]
J Aracena, ´E. Fanchon, M. Montalva, and M. Noual. Combinatorics on update digraphs in Boolean networks.Discrete Applied Mathematics, 159:401–409, 2011
work page 2011
-
[5]
J. Aracena, E. Goles, A. Moreira, and L. Salinas. On the robustness of update schedules in Boolean networks.Biosystems, 97:1–8, 2009
work page 2009
-
[6]
N. P. Bhatia and G. P. Szeg¨ o.Dynamical Systems: Stability Theory and Applications, volume 35 ofLecture Notes in Mathematics. Springer, Berlin, 1967
work page 1967
-
[7]
G. D. Birkhoff.Dynamical Systems, volume 9 ofAMS Colloquium Publications. American Mathematical Society, New York, 1927
work page 1927
-
[8]
Bowen.ω-limit sets for axiom A diffeomorphisms.Journal of Differential Equations, 18:333–339, 1975
R. Bowen.ω-limit sets for axiom A diffeomorphisms.Journal of Differential Equations, 18:333–339, 1975
work page 1975
-
[9]
C. Conley.Isolated invariant sets and the Morse index, volume 38 ofRegional Conference Series in Mathematics. American Mathematical Society, Providence, 1978
work page 1978
-
[10]
T. H. Cormen, C. E. Leiserson, R. L. Rivest, and C. Stein.Introduction to Algorithms. MIT Press and McGraw-Hill, Cambridge, 2nd edition, 2001
work page 2001
-
[11]
M. Cosnard and J. Demongeot. Attracteurs : une approche d´ eterministe.Comptes rendus de l’Acad´ emie des sciences. S´ erie 1, Math´ ematique, 300:551–556, 1985
work page 1985
-
[12]
Demongeot.Seven things I know about them, volume 42 ofEmergence, Complexity and Computation
J. Demongeot.Seven things I know about them, volume 42 ofEmergence, Complexity and Computation. Springer, Cham, 2022
work page 2022
-
[13]
J. Demongeot, A. Elena, and S. Sen´ e. Robustness in regulatory networks: a multi- disciplinary approach.Acta Biotheoretica, 56:27–49, 2008
work page 2008
-
[14]
J. Demongeot and S. Sen´ e. About block-parallel Boolean networks: a position paper. Natural Computing, 19:5–13, 2020
work page 2020
-
[15]
I. Donoso-Leiva, E. Goles, M. R´ ıos-Wilson, and S. Sen´ e. Impact of (a)synchronism on ECA: Towards a new classification.Chaos, Solitons & Fractals, 199:116601, 2025
work page 2025
-
[16]
L. Garrido and C. Sim´ o. Some ideas about strange attractors. InDynamical Systems and Chaos, volume 179 ofLecture Notes in Physics, pages 1–28, Berlin, 1983. Springer. 24
work page 1983
-
[17]
E. Goles and S. Mart´ ınez.Neural and Automata Networks: Dynamical Behavior and Ap- plications. Mathematics and Its Application. Kluwer Academic Publishers, 1990
work page 1990
- [18]
- [19]
-
[20]
E. Goles and J. Olivos. Periodic behaviour of generalized threshold functions.Discrete Mathematics, 30:187–189, 1980
work page 1980
-
[21]
J. Guckenheimer and P. Holmes.Nonlinear Oscillations, Dynamical Systems, and Bifurca- tions of Vector Fields, volume 42 ofApplied Mathematical Sciences. Springer, New York, 1983
work page 1983
-
[22]
I. Harvey and T. Bossomaier. Time out of joint, attractors in asynchronous random Boolean networks. InECAL 1997: Fourth European Conference on Artificial Life, pages 67–75, Cambridge, 1997. MIT Press
work page 1997
-
[23]
J. J. Hopfield. Neural networks and physical systems with emergent collective computa- tional abilities.Proceedings of the National Academy od Sciences of the United States of America, 79:2554–2558, 1982
work page 1982
-
[24]
S. A. Kauffman. Metabolic stability and epigenesis in randomly constructed genetic nets. Journal of Theoretical Biology, 22:437–467, 1969
work page 1969
-
[25]
S. C. Kleene.Representation of events in nerve nets and finite automata. Princeton Uni- versity Press, Princeton, 1951
work page 1951
-
[26]
G.-L. Leclerc de Buffon.Histoire naturelle, g´ en´ erale et particuli` ere, avec la description du Cabinet du Roy. Imprimerie Royale, Paris, 1749
-
[27]
C. F. Mayo and C. E. Roberts. Sequential, parallel and vector solution of ordinary differ- ential equations on a hypercube.Computers & Mathematics with Applications, 18:797–808, 1989
work page 1989
-
[28]
W. S. McCulloch and W. Pitts. A logical calculus of the ideas immanent in nervous activity. The Bulletin of Mathematical Biophysics, 5:115–133, 1943
work page 1943
-
[29]
L. Paulev´ e and S. Sen´ e.Boolean networks and their dynamics: the impact of updates. Wiley, Hoboken, 2022
work page 2022
- [30]
- [31]
-
[32]
K. Perrot, S. Sen´ e, and L. Tapin. Complexity of Boolean automata networks under block- parallel update modes. In3rd Symposium on Algorithmic Foundations of Dynamic Networks (SAND 2024), volume 292 ofLeibniz International Proceedings in Informatics, pages 19:1– 19:19, Dagstuhl, 2024. Schloss Dagstuhl – Leibniz-Zentrum f¨ ur Informatik. 25
work page 2024
- [33]
- [34]
-
[35]
F. Robert.Discrete Iterations: A Metric Study, volume 6 ofSpringer Series in Computa- tional Mathematics. Springer, 1986
work page 1986
-
[36]
D. Ruelle. Small random perturbations of dynamical systems and the definition of attrac- tors.Communications in Mathematical Physics, 82:137–151, 1981
work page 1981
-
[37]
E. L. P. Ruivo, P. P. Balbi, M. Montalva-Medel, and K. Perrot. Maximum sensitivity to update schedules of elementary cellular automata over infinite configurations.Information and Computation, 274:104538, 2020
work page 2020
-
[38]
Y. G. Sinai. The stochasticity of dynamical systems.Selecta Mathematica Sovietica, 1:100– 119, 1981
work page 1981
-
[39]
S. Smale. Differentiable dynamical systems.Bulletin of the American Mathematical Society, 73:747–817, 1967
work page 1967
-
[40]
R. E. Tarjan. Depth-first seach and linear graph algorithms.SIAM Journal on Computing, 1:146–160, 1972
work page 1972
-
[41]
Thom.Stabilit´ e structurelle et morphogen` ese
R. Thom.Stabilit´ e structurelle et morphogen` ese. InterEditions, Paris, 1972
work page 1972
-
[42]
R. Thomas. Boolean formalization of genetic control circuits.Journal of Theoretical Biology, 42:563–585, 1973
work page 1973
-
[43]
R. Tomassone, C. Dervin, and J.-P. Masson.Biom´ etrie : Mod´ elisation de ph´ enom` enes biologiques. Masson, Paris, 1993
work page 1993
-
[44]
D. Vincent, F. Parsopoulou, L. Martin, C. Gaboriaud, J. Demongeot, G. Loules, S. Fischer, S. Cichon, A. E. Germenis, A. Ghannam, and C. Drouet. Hereditary angioedema with normal C1 inhibitor associated with carboxypeptidase N deficiency.Journal of Allergy and Clinical Immunology: Global, 3:100223, 2024
work page 2024
-
[45]
R. F. Williams. Expanding attractors.Publications Math´ ematiques de l’Institut des Hautes ´Etudes Scientifiques, 43:169–203, 1974
work page 1974
-
[46]
S. Zhang, P. Huang, H. Dai, Q. Li, L. Hu, J. Peng, S. Jiang, Y. Xu, Z. Wu, H. Nie, Z. Zhang, W. Yin, X. Zhang, and J. Lu. TIMELESS regulates sphingolipid metabolism and tumor cell growth through Sp1/ACER2/S1P axis in ER-positive breast cancer.Cell Death & Disease, 11:892, 2020. 26
work page 2020
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.