Mostly-monocular responses and other visual functions in a multiscale network model of Macaque V1
Pith reviewed 2026-06-26 12:19 UTC · model grok-4.3
The pith
In a multiscale model of macaque V1, narrow binocular strips emerge along ocular dominance column borders when 10-30% of interactions near boundaries are cross-columnar, and layer 6 feedback is largely monocular.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
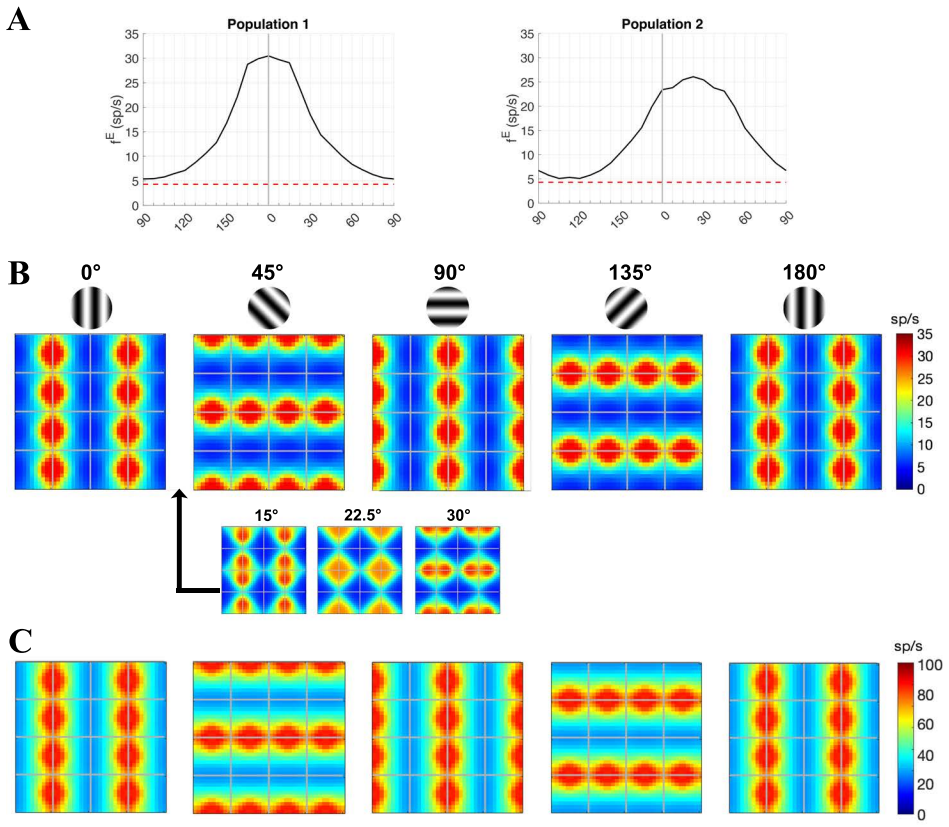
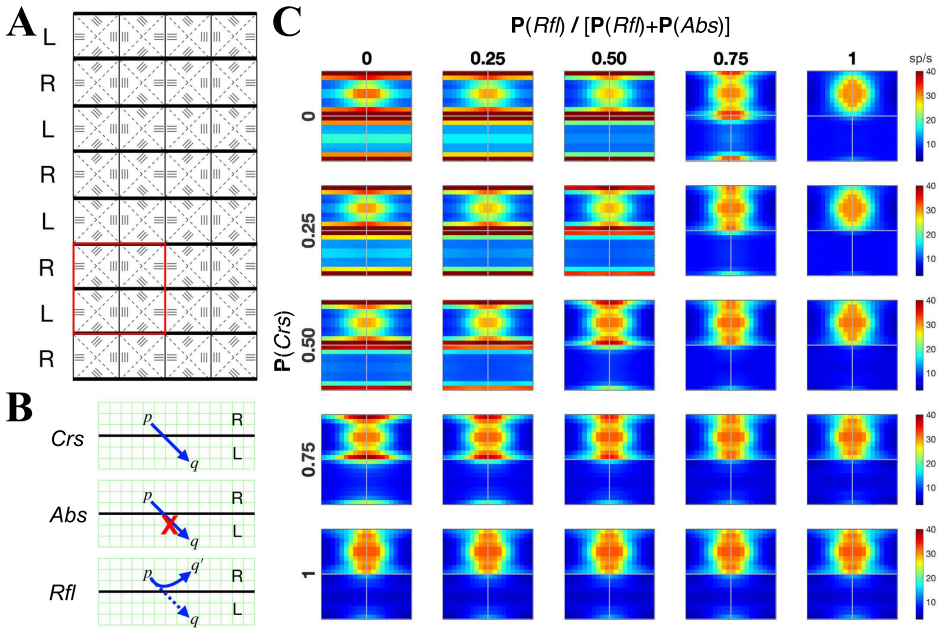
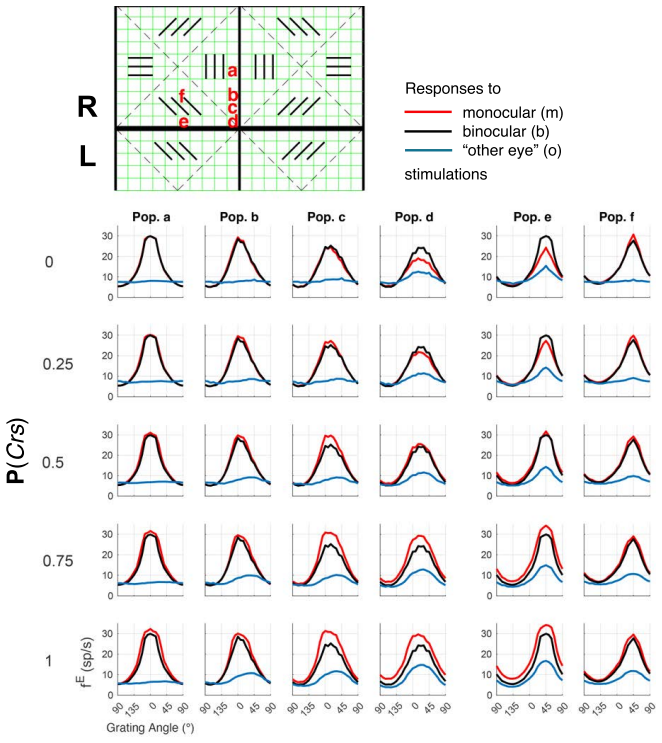
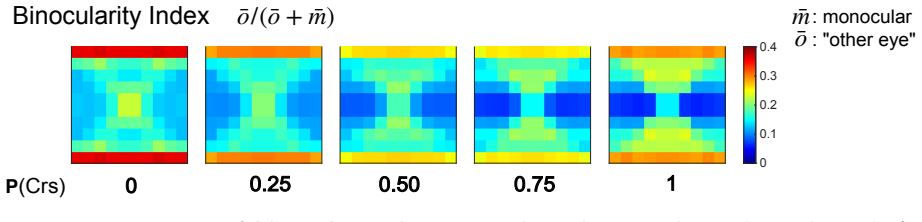
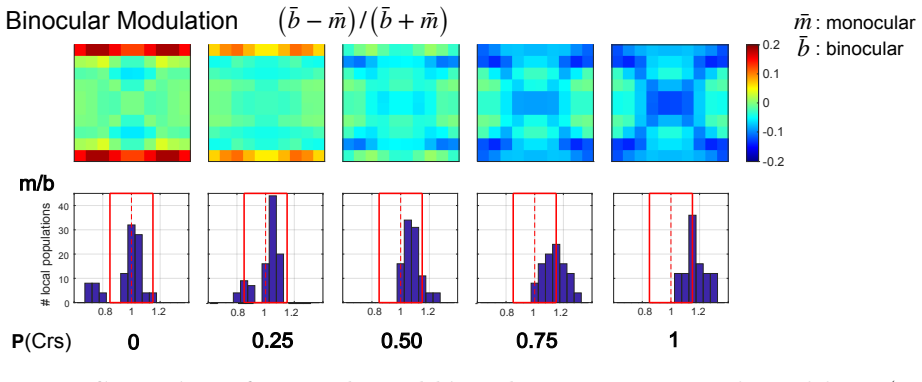
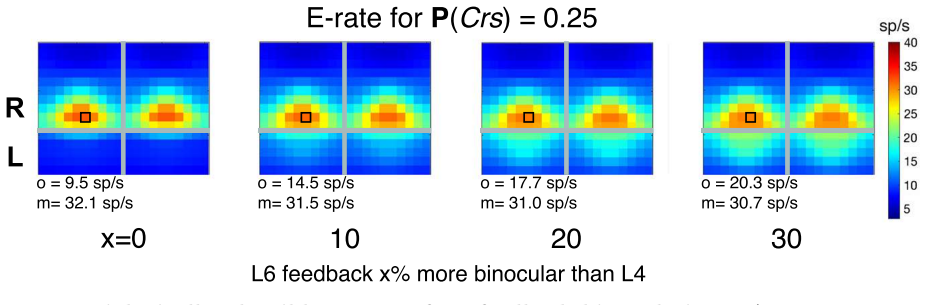
Using a multiscale network model of macaque V1, narrow binocular strips emerge along the borders of ocular dominance columns in layer 4Cα, consistent with experiments particularly when 10-30% of interactions near ODC boundaries are cross-columnar, and feedback from layer 6 is largely monocular, allowing inference of the neuroanatomical origins of binocular response.
What carries the argument
The multiscale network model approximating detailed V1 circuitry to test hypotheses on ocular dominance and binocularity in layer 4Cα.
If this is right
- Narrow binocular strips form along ocular dominance column borders.
- 10-30% cross-columnar interactions near boundaries best match experimental data.
- Feedback from layer 6 remains largely monocular.
- Multiscale modeling can bridge anatomy and function in V1.
Where Pith is reading between the lines
- The border effect may generalize to other types of cortical columns or sensory modalities.
- Varying the interaction fraction in the model could predict how wiring changes affect binocularity.
- The approach might be used to study gradual integration in higher visual areas.
Load-bearing premise
The multiscale approximation and the 10-30% cross-columnar interaction fraction near ODC boundaries are sufficient to capture the dominant anatomical drivers of binocularity without missing critical unmodeled factors.
What would settle it
Direct measurements in macaque V1 showing either no narrow binocular strips along ODC borders or a cross-columnar interaction fraction near boundaries outside the 10-30% range would falsify the model's main results.
Figures

read the original abstract
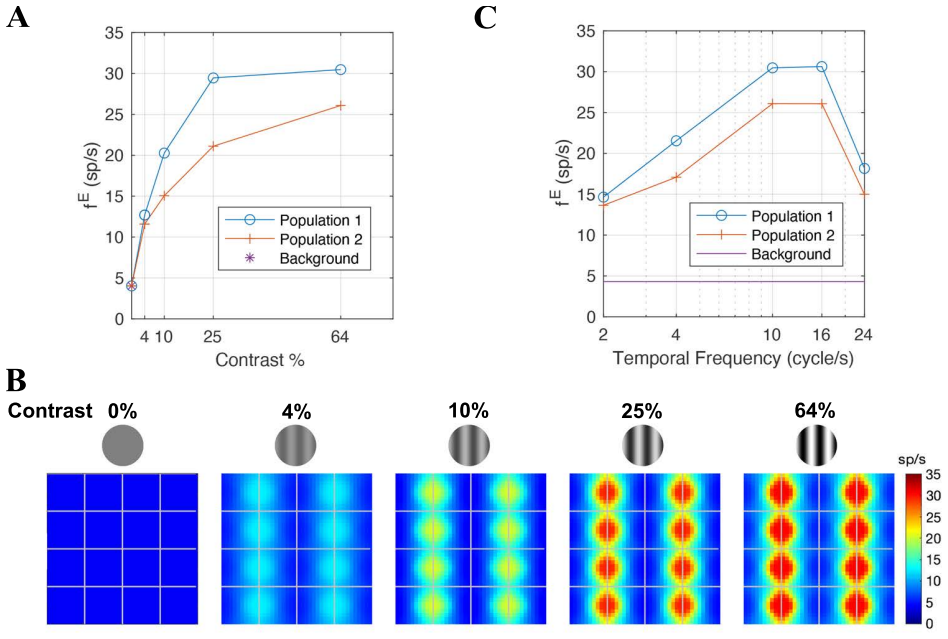
Visual signals from the two eyes merge gradually as they pass through the primary visual cortex (V1). Here we use a computational model of Macaque V1 to study the first stage of this integration along the magnocellular pathway, in layer 4C$\alpha$, aiming to infer neuroanatomical origins of binocular response. It is known that neurons in layer 4C$\alpha$ are predominantly monocular, though some do exhibit varying degrees of binocularity. We find (1) the emergence of narrow binocular strips along borders of ocular dominance columns (ODC), a finding that aligns with experiments; (2) most consistent with data is when $10-30\%$ of interactions near ODC boundaries are cross-columnar; and (3) feedback from layer 6 is largely monocular. These results were obtained through systematic hypothesis testing using a multiscale model that is orders of magnitude faster than its biologically-detailed predecessors. We propose that multiscale modeling can be an effective tool for bridging anatomy and function.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript presents a multiscale network model of Macaque V1 layer 4Cα to study the first stage of binocular integration along the magnocellular pathway. It reports three main findings obtained via systematic hypothesis testing: (1) emergence of narrow binocular strips along ocular dominance column (ODC) borders that align with experiments; (2) best consistency with data when 10-30% of interactions near ODC boundaries are cross-columnar; and (3) largely monocular feedback from layer 6. The model is described as orders of magnitude faster than detailed predecessors.
Significance. If the multiscale approximation is shown to faithfully capture local connectivity near ODC borders, the work offers an efficient computational tool for linking neuroanatomy to visual function and for systematic exploration of parameter regimes in V1 models. The emphasis on hypothesis testing and the identification of a narrow range of cross-columnar interactions provide a concrete, falsifiable link between anatomy and the observed mostly-monocular responses.
major comments (2)
- [Abstract (model description) and hypothesis-testing results] The central claims on binocular-strip emergence and the 10-30% cross-columnar fraction rest on the multiscale approximation's ability to represent local border interactions without altering effective cross-eye synaptic drive; no control recomputation of the same observables in a non-multiscale or finer-scale limit is reported, leaving open whether the strips and percentage range are robust or artifacts of coarse-graining.
- [Abstract and results on cross-columnar interactions] The 10-30% range is stated as 'most consistent with data,' yet the manuscript provides no explicit statement on whether this interval was pre-specified before running the simulations or selected after inspecting model outputs; this directly affects the strength of the claim that the model aligns with experimental binocularity patterns.
minor comments (1)
- [Abstract] The abstract refers to 'systematic hypothesis testing' and 'alternative parameter regimes' but the provided text does not detail the full exploration of error bars or the precise definition of the observables used to score consistency with data.
Simulated Author's Rebuttal
We thank the referee for their constructive comments on our manuscript. We address the major comments point by point below, and will incorporate clarifications in the revised version where appropriate.
read point-by-point responses
-
Referee: [Abstract (model description) and hypothesis-testing results] The central claims on binocular-strip emergence and the 10-30% cross-columnar fraction rest on the multiscale approximation's ability to represent local border interactions without altering effective cross-eye synaptic drive; no control recomputation of the same observables in a non-multiscale or finer-scale limit is reported, leaving open whether the strips and percentage range are robust or artifacts of coarse-graining.
Authors: The multiscale approximation is designed to faithfully represent local connectivity near ODC borders by scaling interactions appropriately, thereby preserving the effective cross-eye synaptic drive. We did not include a non-multiscale control because the detailed model is computationally infeasible for the systematic parameter exploration performed here. However, we will add a new subsection in the methods or discussion to elaborate on the theoretical justification for the approximation and why it is unlikely to introduce artifacts in the reported observables. This addresses the concern without requiring infeasible recomputations. revision: yes
-
Referee: [Abstract and results on cross-columnar interactions] The 10-30% range is stated as 'most consistent with data,' yet the manuscript provides no explicit statement on whether this interval was pre-specified before running the simulations or selected after inspecting model outputs; this directly affects the strength of the claim that the model aligns with experimental binocularity patterns.
Authors: We agree that transparency regarding the determination of the 10-30% range is important. This range was identified through systematic hypothesis testing by varying the cross-columnar interaction percentage and comparing the resulting response patterns to experimental data. It was not pre-specified but emerged from the exploration. In the revised manuscript, we will explicitly state in the results section how the range was determined and note that it is the outcome of the hypothesis-testing procedure rather than an a priori prediction. revision: yes
Circularity Check
Cross-columnar interaction fraction tuned to match binocular data; emergence claim otherwise anatomy-driven
specific steps
-
fitted input called prediction
[Abstract]
"most consistent with data is when 10-30% of interactions near ODC boundaries are cross-columnar"
The model systematically varies the cross-columnar interaction fraction near ODC boundaries and reports the 10-30% interval as the range most consistent with experimental binocular-response data; the reported interval is therefore the direct output of the fitting procedure rather than an independent derivation from first-principles connectivity.
full rationale
The paper's central outputs are (1) emergence of narrow binocular strips at ODC borders as a model consequence of anatomical connectivity rules and (2) identification of the 10-30% cross-columnar fraction near boundaries as most consistent with data. The second is obtained by systematic variation of that fraction inside the multiscale model and direct comparison to experimental binocularity measurements; this is a fitted-input step rather than an a-priori prediction. No self-citation chain, uniqueness theorem, or ansatz smuggling is visible in the provided text that would render the strip-emergence result tautological. The multiscale approximation is presented as a computational tool whose validity is assumed rather than re-derived here. The derivation therefore retains independent anatomical content but carries moderate circularity burden on the quantitative percentage claim.
Axiom & Free-Parameter Ledger
free parameters (1)
- fraction of cross-columnar interactions near ODC boundaries
axioms (1)
- domain assumption The multiscale network reduction accurately represents the dominant local connectivity rules in layer 4Cα.
Reference graph
Works this paper leans on
-
[1]
Theoretical neuroscience rising.Neuron, 60(3):489–495, 2008
Larry F Abbott. Theoretical neuroscience rising.Neuron, 60(3):489–495, 2008
2008
-
[2]
Theory and simulation in neuro- science.Science, 338(6103):60–65, 2012
Wulfram Gerstner, Henning Sprekeler, and Gustavo Deco. Theory and simulation in neuro- science.Science, 338(6103):60–65, 2012. 19
2012
-
[3]
Computational neuroscience: beyond the local circuit.Current opinion in neurobiology, 25:xiii–xviii, 2014
Haim Sompolinsky. Computational neuroscience: beyond the local circuit.Current opinion in neurobiology, 25:xiii–xviii, 2014
2014
-
[4]
The scientific case for brain simulations.Neuron, 102(4):735–744, 2019
Gaute T Einevoll, Alain Destexhe, Markus Diesmann, Sonja Gr¨ un, Viktor Jirsa, Marc de Kamps, Michele Migliore, Torbjørn V Ness, Hans E Plesser, and Felix Sch¨ urmann. The scientific case for brain simulations.Neuron, 102(4):735–744, 2019
2019
-
[5]
Modeling single-neuron dynamics and computations: a balance of detail and abstraction.Science, 314(5796):80–85, 2006
Andreas VM Herz, Tim Gollisch, Christian K Machens, and Dieter Jaeger. Modeling single-neuron dynamics and computations: a balance of detail and abstraction.Science, 314(5796):80–85, 2006
2006
-
[6]
On the nature and use of models in network neuroscience.Nature Reviews Neuroscience, 19(9):566–578, 2018
Danielle S Bassett, Perry Zurn, and Joshua I Gold. On the nature and use of models in network neuroscience.Nature Reviews Neuroscience, 19(9):566–578, 2018
2018
-
[7]
The cell-type specific cortical microcircuit: relating structure and activity in a full-scale spiking network model.Cerebral cortex, 24(3):785–806, 2014
Tobias C Potjans and Markus Diesmann. The cell-type specific cortical microcircuit: relating structure and activity in a full-scale spiking network model.Cerebral cortex, 24(3):785–806, 2014
2014
-
[8]
Using goal-driven deep learning models to understand sensory cortex.Nature neuroscience, 19(3):356–365, 2016
Daniel LK Yamins and James J DiCarlo. Using goal-driven deep learning models to understand sensory cortex.Nature neuroscience, 19(3):356–365, 2016
2016
-
[9]
Systematic integration of structural and functional data into multi-scale models of mouse primary visual cortex.Neuron, 106(3):388–403, 2020
Yazan N Billeh, Binghuang Cai, Sergey L Gratiy, Kael Dai, Ramakrishnan Iyer, Nathan W Gouwens, Reza Abbasi-Asl, Xiaoxuan Jia, Joshua H Siegle, Shawn R Olsen, et al. Systematic integration of structural and functional data into multi-scale models of mouse primary visual cortex.Neuron, 106(3):388–403, 2020
2020
-
[10]
Combining hypothesis-and data-driven neuroscience modeling in fair work- flows.Elife, 11:e69013, 2022
Olivia Eriksson, Upinder Singh Bhalla, Kim T Blackwell, Sharon M Crook, Daniel Keller, Andrei Kramer, Marja-Leena Linne, Ausra Saudargien˙ e, Rebecca C Wade, and Jeanette Hell- gren Kotaleski. Combining hypothesis-and data-driven neuroscience modeling in fair work- flows.Elife, 11:e69013, 2022
2022
-
[11]
A data-based large-scale model for pri- mary visual cortex enables brain-like robust and versatile visual processing.Science Advances, 8(44):eabq7592, 2022
Guozhang Chen, Franz Scherr, and Wolfgang Maass. A data-based large-scale model for pri- mary visual cortex enables brain-like robust and versatile visual processing.Science Advances, 8(44):eabq7592, 2022
2022
-
[12]
Large-scale mechanistic models of brain circuits with biophysically and morphologically detailed neurons.Journal of Neuroscience, 44(40), 2024
Salvador Dura-Bernal, Beatriz Herrera, Carmen Lupascu, Brianna M Marsh, Daniela Gandolfi, Addolorata Marasco, Samuel Neymotin, Armando Romani, Sergio Solinas, Maxim Bazhenov, et al. Large-scale mechanistic models of brain circuits with biophysically and morphologically detailed neurons.Journal of Neuroscience, 44(40), 2024
2024
-
[13]
Mapping effective connectivity by virtually perturbing a surrogate brain.Nature Methods, 22(6):1376–1385, 2025
Zixiang Luo, Kaining Peng, Zhichao Liang, Shengyuan Cai, Chenyu Xu, Dan Li, Yu Hu, Changsong Zhou, and Quanying Liu. Mapping effective connectivity by virtually perturbing a surrogate brain.Nature Methods, 22(6):1376–1385, 2025
2025
-
[14]
Reconstruction and simulation of neocortical microcircuitry.Cell, 163(2):456– 492, 2015
Henry Markram, Eilif Muller, Srikanth Ramaswamy, Michael W Reimann, Marwan Abdellah, Carlos Aguado Sanchez, Anastasia Ailamaki, Lidia Alonso-Nanclares, Nicolas Antille, Selim Arsever, et al. Reconstruction and simulation of neocortical microcircuitry.Cell, 163(2):456– 492, 2015
2015
-
[15]
Efficient models of cortical activity via local dynamic equilibria and coarse-grained interactions.Proceedings of the National Academy of Sciences, 121(27):e2320454121, 2024
Zhuo-Cheng Xiao, Kevin K Lin, and Lai-Sang Young. Efficient models of cortical activity via local dynamic equilibria and coarse-grained interactions.Proceedings of the National Academy of Sciences, 121(27):e2320454121, 2024. 20
2024
-
[16]
Neuroimaging weighs in: humans meet macaques in “primate” visual cortex.Journal of Neuroscience, 23(10):3981–3989, 2003
Roger BH Tootell, Doris Tsao, and Wim Vanduffel. Neuroimaging weighs in: humans meet macaques in “primate” visual cortex.Journal of Neuroscience, 23(10):3981–3989, 2003
2003
-
[17]
Comparative mapping of higher visual areas in monkeys and humans.Trends in cognitive sciences, 8(7):315–324, 2004
Guy A Orban, David Van Essen, and Wim Vanduffel. Comparative mapping of higher visual areas in monkeys and humans.Trends in cognitive sciences, 8(7):315–324, 2004
2004
-
[18]
Parallel colour-opponent pathways to primary visual cortex.Nature, 426(6967):668–671, 2003
Soumya Chatterjee and Edward M Callaway. Parallel colour-opponent pathways to primary visual cortex.Nature, 426(6967):668–671, 2003
2003
-
[19]
Parallel processing strategies of the primate visual system.Nature reviews neuroscience, 10(5):360–372, 2009
Jonathan J Nassi and Edward M Callaway. Parallel processing strategies of the primate visual system.Nature reviews neuroscience, 10(5):360–372, 2009
2009
-
[20]
Orientation selectivity from very sparse lgn inputs in a comprehensive model of macaque v1 cortex.Journal of Neuroscience, 36(49):12368–12384, 2016
Logan Chariker, Robert Shapley, and Lai-Sang Young. Orientation selectivity from very sparse lgn inputs in a comprehensive model of macaque v1 cortex.Journal of Neuroscience, 36(49):12368–12384, 2016
2016
-
[21]
Contrast response in a comprehensive network model of macaque v1.Journal of vision, 20(4):16–16, 2020
Logan Chariker, Robert Shapley, and Lai-Sang Young. Contrast response in a comprehensive network model of macaque v1.Journal of vision, 20(4):16–16, 2020
2020
-
[22]
A computational model of direction selectivity in macaque v1 cortex based on dynamic differences between on and off pathways.Journal of Neuroscience, 42(16):3365–3380, 2022
Logan Chariker, Robert Shapley, Michael Hawken, and Lai-Sang Young. A computational model of direction selectivity in macaque v1 cortex based on dynamic differences between on and off pathways.Journal of Neuroscience, 42(16):3365–3380, 2022
2022
-
[23]
Receptive fields and functional architecture of monkey striate cortex.The Journal of physiology, 195(1):215–243, 1968
David H Hubel and Torsten N Wiesel. Receptive fields and functional architecture of monkey striate cortex.The Journal of physiology, 195(1):215–243, 1968
1968
-
[24]
Orientation selectivity in macaque v1: diversity and laminar dependence.Journal of neuroscience, 22(13):5639–5651, 2002
Dario L Ringach, Robert M Shapley, and Michael J Hawken. Orientation selectivity in macaque v1: diversity and laminar dependence.Journal of neuroscience, 22(13):5639–5651, 2002
2002
-
[25]
Area v1.Scholarpedia, 7(7):12105, 2012
Matteo Carandini. Area v1.Scholarpedia, 7(7):12105, 2012
2012
-
[26]
Mechanisms of neuronal computation in mammalian visual cortex.Neuron, 75(2):194–208, 2012
Nicholas J Priebe and David Ferster. Mechanisms of neuronal computation in mammalian visual cortex.Neuron, 75(2):194–208, 2012
2012
-
[27]
Functional anatomy of macaque striate cortex
Roger BH Tootell, Steven L Hamilton, Michael S Silverman, and Eugene Switkes. Functional anatomy of macaque striate cortex. I. ocular dominance, binocular interactions, and baseline conditions.Journal of Neuroscience, 8(5):1500–1530, 1988
1988
-
[28]
Ocular dominance columns: enigmas and challenges
Daniel L Adams and Jonathan C Horton. Ocular dominance columns: enigmas and challenges. The Neuroscientist, 15(1):62–77, 2009
2009
-
[29]
Contrast sensitivity and orientation selectivity in lamina iv of the striate cortex of old world monkeys.Experimental Brain Research, 54(2):367–372, 1984
MJ Hawken and AJ Parker. Contrast sensitivity and orientation selectivity in lamina iv of the striate cortex of old world monkeys.Experimental Brain Research, 54(2):367–372, 1984
1984
-
[30]
Local circuits and ocular domi- nance columns in monkey striate cortex.Journal of Neuroscience, 9(4):1389–1399, 1989
Lawrence C Katz, Charles D Gilbert, and Torsten N Wiesel. Local circuits and ocular domi- nance columns in monkey striate cortex.Journal of Neuroscience, 9(4):1389–1399, 1989
1989
-
[31]
Alessandra Angelucci and Paul C Bressloff. Contribution of feedforward, lateral and feedback connections to the classical receptive field center and extra-classical receptive field surround of primate v1 neurons.Progress in brain research, 154:93–120, 2006
2006
-
[32]
Binocular modulation of monocular v1 neurons.Current Biology, 29(3):381–391, 2019
Kacie Dougherty, Michele A Cox, Jacob A Westerberg, and Alexander Maier. Binocular modulation of monocular v1 neurons.Current Biology, 29(3):381–391, 2019. 21
2019
-
[33]
Binocular integration in the primate primary visual cortex.Annual Review of Vision Science, 8(1):345–360, 2022
A Maier, MA Cox, JA Westerberg, and K Dougherty. Binocular integration in the primate primary visual cortex.Annual Review of Vision Science, 8(1):345–360, 2022
2022
-
[34]
Contributions of individual layer 6 pyramidal neurons to local circuitry in macaque primary visual cortex.Journal of neuroscience, 16(8):2724–2739, 1996
Anne K Wiser and Edward M Callaway. Contributions of individual layer 6 pyramidal neurons to local circuitry in macaque primary visual cortex.Journal of neuroscience, 16(8):2724–2739, 1996
1996
-
[35]
The circuitry of v1 and v2: integration of color, form, and motion.Annu
Lawrence C Sincich and Jonathan C Horton. The circuitry of v1 and v2: integration of color, form, and motion.Annu. Rev. Neurosci., 28(1):303–326, 2005
2005
-
[36]
Receptive fields, binocular interaction and functional architecture in the cat’s visual cortex.The Journal of physiology, 160(1):106, 1962
David H Hubel and Torsten N Wiesel. Receptive fields, binocular interaction and functional architecture in the cat’s visual cortex.The Journal of physiology, 160(1):106, 1962
1962
-
[37]
Orientation selectivity, preference, and continuity in monkey striate cortex
Gary G Blasdel. Orientation selectivity, preference, and continuity in monkey striate cortex. Journal of Neuroscience, 12(8):3139–3161, 1992
1992
-
[38]
Geometry of orientation and ocular dominance columns in monkey striate cortex.Journal of Neuroscience, 13(10):4114–4129, 1993
Klaus Obermayer and Gary G Blasdel. Geometry of orientation and ocular dominance columns in monkey striate cortex.Journal of Neuroscience, 13(10):4114–4129, 1993
1993
-
[39]
Anatomical substrates for functional columns in macaque monkey primary visual cortex.Cerebral cortex, 13(1):15–24, 2003
Jennifer S Lund, Alessandra Angelucci, and Paul C Bressloff. Anatomical substrates for functional columns in macaque monkey primary visual cortex.Cerebral cortex, 13(1):15–24, 2003
2003
-
[40]
Local circuits in primary visual cortex of the macaque monkey.Annual review of neuroscience, 21(1):47–74, 1998
Edward M Callaway. Local circuits in primary visual cortex of the macaque monkey.Annual review of neuroscience, 21(1):47–74, 1998
1998
-
[41]
Orientation selectivity of thalamic input to simple cells of cat visual cortex.Nature, 380(6571):249–252, 1996
David Ferster, Sooyoung Chung, and Heidi Wheat. Orientation selectivity of thalamic input to simple cells of cat visual cortex.Nature, 380(6571):249–252, 1996
1996
-
[42]
Harvey A Swadlow. Efferent neurons and suspected interneurons in binocular visual cortex of the awake rabbit: receptive fields and binocular properties.Journal of neurophysiology, 59(4):1162–1187, 1988
1988
-
[43]
Stimulus feature selectivity in excita- tory and inhibitory neurons in primary visual cortex.Journal of Neuroscience, 27(39):10333– 10344, 2007
Jessica A Cardin, Larry A Palmer, and Diego Contreras. Stimulus feature selectivity in excita- tory and inhibitory neurons in primary visual cortex.Journal of Neuroscience, 27(39):10333– 10344, 2007
2007
-
[44]
Striate cortex of monkey and cat: contrast response function.Journal of neurophysiology, 48(1):217–237, 1982
Duane G Albrecht and David B Hamilton. Striate cortex of monkey and cat: contrast response function.Journal of neurophysiology, 48(1):217–237, 1982
1982
-
[45]
Functional characterization of the extraclassical receptive field in macaque v1: con- trast, orientation, and temporal dynamics.Journal of Neuroscience, 33(14):6230–6242, 2013
Christopher A Henry, Siddhartha Joshi, Dajun Xing, Robert M Shapley, and Michael J Hawken. Functional characterization of the extraclassical receptive field in macaque v1: con- trast, orientation, and temporal dynamics.Journal of Neuroscience, 33(14):6230–6242, 2013
2013
-
[46]
Recurrent excitation in neocortical circuits.Science, 269(5226):981–985, 1995
Rodney J Douglas, Christof Koch, Misha Mahowald, Kevan AC Martin, and Humbert H Suarez. Recurrent excitation in neocortical circuits.Science, 269(5226):981–985, 1995
1995
-
[47]
Contrast invariance of functional maps in cat primary visual cortex.Journal of vision, 4(3):1–1, 2004
Matteo Carandini and Frank Sengpiel. Contrast invariance of functional maps in cat primary visual cortex.Journal of vision, 4(3):1–1, 2004
2004
-
[48]
Temporal-frequency selectivity in monkey visual cortex.Visual neuroscience, 13(3):477–492, 1996
MJ Hawken, Robert M Shapley, and DH Grosof. Temporal-frequency selectivity in monkey visual cortex.Visual neuroscience, 13(3):477–492, 1996. 22
1996
-
[49]
Michael Connolly and David Van Essen. The representation of the visual field in parvicellular and magnocellular layers of the lateral geniculate nucleus in the macaque monkey.Journal of Comparative Neurology, 226(4):544–564, 1984
1984
-
[50]
Structure and function of parallel pathways in the primate early visual system.The Journal of physiology, 566(1):13–19, 2005
Edward M Callaway. Structure and function of parallel pathways in the primate early visual system.The Journal of physiology, 566(1):13–19, 2005
2005
-
[51]
Jonathan C Horton and Davina R Hocking. Monocular core zones and binocular border strips in primate striate cortex revealed by the contrasting effects of enucleation, eyelid suture, and retinal laser lesions on cytochrome oxidase activity.Journal of Neuroscience, 18(14):5433–5455, 1998
1998
-
[52]
Temporal dynamics of binocular integration in primary visual cortex.Journal of vision, 19(12):13–13, 2019
Michele A Cox, Kacie Dougherty, Jacob A Westerberg, Michelle S Schall, and Alexander Maier. Temporal dynamics of binocular integration in primary visual cortex.Journal of vision, 19(12):13–13, 2019
2019
-
[53]
Ocular dominance columns and local projections of layer 6 pyramidal neurons in macaque primary visual cortex.Visual neuroscience, 14(2):241– 251, 1997
Anne K Wiser and Edward M Callaway. Ocular dominance columns and local projections of layer 6 pyramidal neurons in macaque primary visual cortex.Visual neuroscience, 14(2):241– 251, 1997
1997
-
[54]
The development of ocular domi- nance columns in normal and visually deprived monkeys.Journal of Comparative Neurology, 191(1):1–51, 1980
Simon Le Vay, Torsten N Wiesel, and David H Hubel. The development of ocular domi- nance columns in normal and visually deprived monkeys.Journal of Comparative Neurology, 191(1):1–51, 1980
1980
-
[55]
Ocular dominance and disparity coding in cat visual cortex
Simon LeVay and Thomas Voigt. Ocular dominance and disparity coding in cat visual cortex. Visual neuroscience, 1(4):395–414, 1988
1988
-
[56]
Stimulating both eyes with matching stimuli enhances v1 responses.Iscience, 25(5), 2022
Blake A Mitchell, Kacie Dougherty, Jacob A Westerberg, Brock M Carlson, Lo¨ ıc Daumail, Alexander Maier, and Michele A Cox. Stimulating both eyes with matching stimuli enhances v1 responses.Iscience, 25(5), 2022
2022
-
[57]
Organizing principles of cortical layer 6.Frontiers in neural circuits, 4:750, 2010
Farran Briggs. Organizing principles of cortical layer 6.Frontiers in neural circuits, 4:750, 2010
2010
-
[58]
Neocortical layer 6, a review.Frontiers in neuroanatomy, 4:1339, 2010
Alex M Thomson. Neocortical layer 6, a review.Frontiers in neuroanatomy, 4:1339, 2010
2010
-
[59]
Quantitative studies of single-cell properties in monkey striate cortex
Peter H Schiller, Barbara L Finlay, and Susan F Volman. Quantitative studies of single-cell properties in monkey striate cortex. ii. orientation specificity and ocular dominance.Journal of neurophysiology, 39(6):1320–1333, 1976
1976
-
[60]
David McLaughlin, Robert Shapley, Michael Shelley, and Dingeman J Wielaard. A neuronal network model of macaque primary visual cortex (v1): Orientation selectivity and dynamics in the input layer 4ca.Proceedings of the National Academy of Sciences, 97(14):8087–8092, 2000
2000
-
[61]
Large-scale modeling of the primary visual cortex: influence of cortical architecture upon neuronal response.Journal of Physiology- Paris, 97(2-3):237–252, 2003
David McLaughlin, Robert Shapley, and Michael Shelley. Large-scale modeling of the primary visual cortex: influence of cortical architecture upon neuronal response.Journal of Physiology- Paris, 97(2-3):237–252, 2003
2003
-
[62]
An egalitarian network model for the emergence of simple and complex cells in visual cortex.Proceedings of the National Academy of Sciences, 101(1):366–371, 2004
Louis Tao, Michael Shelley, David McLaughlin, and Robert Shapley. An egalitarian network model for the emergence of simple and complex cells in visual cortex.Proceedings of the National Academy of Sciences, 101(1):366–371, 2004. 23
2004
-
[63]
Heterogeneous multiscale method: a general methodology for multiscale modeling.Physical Review B, 67(9):092101, 2003
E Weinan, Bjorn Engquist, and Zhongyi Huang. Heterogeneous multiscale method: a general methodology for multiscale modeling.Physical Review B, 67(9):092101, 2003
2003
-
[64]
Equation-free, coarse-grained multiscale computa- tion: enabling microscopic simulators to perform system-level analysis.Commun
Ioannis G Kevrekidis, C William Gear, James M Hyman, Panagiotis G Kevrekidis, Olof Run- borg, Constantinos Theodoropoulos, et al. Equation-free, coarse-grained multiscale computa- tion: enabling microscopic simulators to perform system-level analysis.Commun. Math. Sci, 1(4):715–762, 2003
2003
-
[65]
The heterogeneous multiscale method.Acta Numerica, 21:1–87, 2012
Assyr Abdulle, E Weinan, Bj¨ orn Engquist, and Eric Vanden-Eijnden. The heterogeneous multiscale method.Acta Numerica, 21:1–87, 2012
2012
-
[66]
Netpyne, a tool for data-driven multiscale modeling of brain circuits.Elife, 8:e44494, 2019
Salvador Dura-Bernal, Benjamin A Suter, Padraig Gleeson, Matteo Cantarelli, Adrian Quin- tana, Facundo Rodriguez, David J Kedziora, George L Chadderdon, Cliff C Kerr, Samuel A Neymotin, et al. Netpyne, a tool for data-driven multiscale modeling of brain circuits.Elife, 8:e44494, 2019
2019
-
[67]
Colloquium: Multiscale modeling of brain network organization.Reviews of Modern Physics, 94(3):031002, 2022
Charley Presigny and Fabrizio De Vico Fallani. Colloquium: Multiscale modeling of brain network organization.Reviews of Modern Physics, 94(3):031002, 2022
2022
-
[68]
The quest for multiscale brain modeling.Trends in neuro- sciences, 45(10):777–790, 2022
Egidio D’Angelo and Viktor Jirsa. The quest for multiscale brain modeling.Trends in neuro- sciences, 45(10):777–790, 2022
2022
-
[69]
Descriptive multiscale modeling in data-driven neuroscience.Synthese, 200(2):129, 2022
Philipp Haueis. Descriptive multiscale modeling in data-driven neuroscience.Synthese, 200(2):129, 2022
2022
-
[70]
Anatomy of hierarchy: feedforward and feedback pathways in macaque visual cortex.Journal of compara- tive neurology, 522(1):225–259, 2014
Nikola T Markov, Julien Vezoli, Pascal Chameau, Arnaud Falchier, Ren´ e Quilodran, Cyril Huissoud, Camille Lamy, Pierre Misery, Pascale Giroud, Shimon Ullman, et al. Anatomy of hierarchy: feedforward and feedback pathways in macaque visual cortex.Journal of compara- tive neurology, 522(1):225–259, 2014
2014
-
[71]
The effects of contrast on visual orientation and spatial frequency discrimination: a comparison of single cells and behavior.Journal of neurophysiology, 57(3):773–786, 1987
Bernt C Skottun, Arthur Bradley, Gary Sclar, Izumi Ohzawa, and Ralph D Freeman. The effects of contrast on visual orientation and spatial frequency discrimination: a comparison of single cells and behavior.Journal of neurophysiology, 57(3):773–786, 1987
1987
-
[72]
Circuits and mechanisms for surround modulation in visual cortex
Alessandra Angelucci, Maryam Bijanzadeh, Lauri Nurminen, Frederick Federer, Sam Merlin, and Paul C Bressloff. Circuits and mechanisms for surround modulation in visual cortex. Annual review of neuroscience, 40(1):425–451, 2017
2017
-
[73]
Laminar subnetworks of response suppression in macaque primary visual cortex.Journal of Neuroscience, 40(39):7436–7450, 2020
Tian Wang, Yang Li, Guanzhong Yang, Weifeng Dai, Yi Yang, Chuanliang Han, Xingyun Wang, Yange Zhang, and Dajun Xing. Laminar subnetworks of response suppression in macaque primary visual cortex.Journal of Neuroscience, 40(39):7436–7450, 2020. 24
2020
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.