On a Generalized Compartment Model for Ethanol Metabolism in the Human Body
Pith reviewed 2026-06-30 03:56 UTC · model grok-4.3
The pith
Generalized compartment model for ethanol metabolism proves global asymptotic stability of its unique equilibrium using a quadratic Lyapunov function.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
The generalized continuous-time compartment model with nonlinear ethanol metabolism rate functions has positive and bounded solutions and possesses a unique equilibrium that is globally asymptotically stable, as shown by constructing an appropriate quadratic Lyapunov function whose derivative is negative definite along trajectories. The corresponding discrete-time model reproduces positivity, boundedness, and global asymptotic stability whenever the time step size satisfies a derived restriction.
What carries the argument
The general class of nonlinear rate functions for hepatic ethanol metabolism, which replaces the Michaelis-Menten form and permits construction of a quadratic Lyapunov function proving global stability.
If this is right
- Solutions remain positive and bounded for nonnegative initial conditions.
- The unique equilibrium is globally asymptotically stable under the stated conditions on the rate functions.
- The discrete-time model inherits positivity, boundedness, and global stability when the time step is sufficiently small.
- Numerical simulations with varied nonlinear rate functions confirm convergence to the equilibrium.
Where Pith is reading between the lines
- The same Lyapunov construction may apply to other compartment models whose elimination rates obey comparable growth bounds.
- The time-step restriction supplies a practical rule for choosing simulation step sizes that preserve qualitative behavior.
Load-bearing premise
The nonlinear rate functions must satisfy positivity, continuity, and suitable monotonicity or growth conditions so that a unique equilibrium exists and a quadratic Lyapunov function can be built with negative definite derivative.
What would settle it
A concrete nonlinear rate function satisfying the paper's technical conditions for which numerical integration shows either multiple equilibria or trajectories that fail to converge to the predicted equilibrium.
Figures

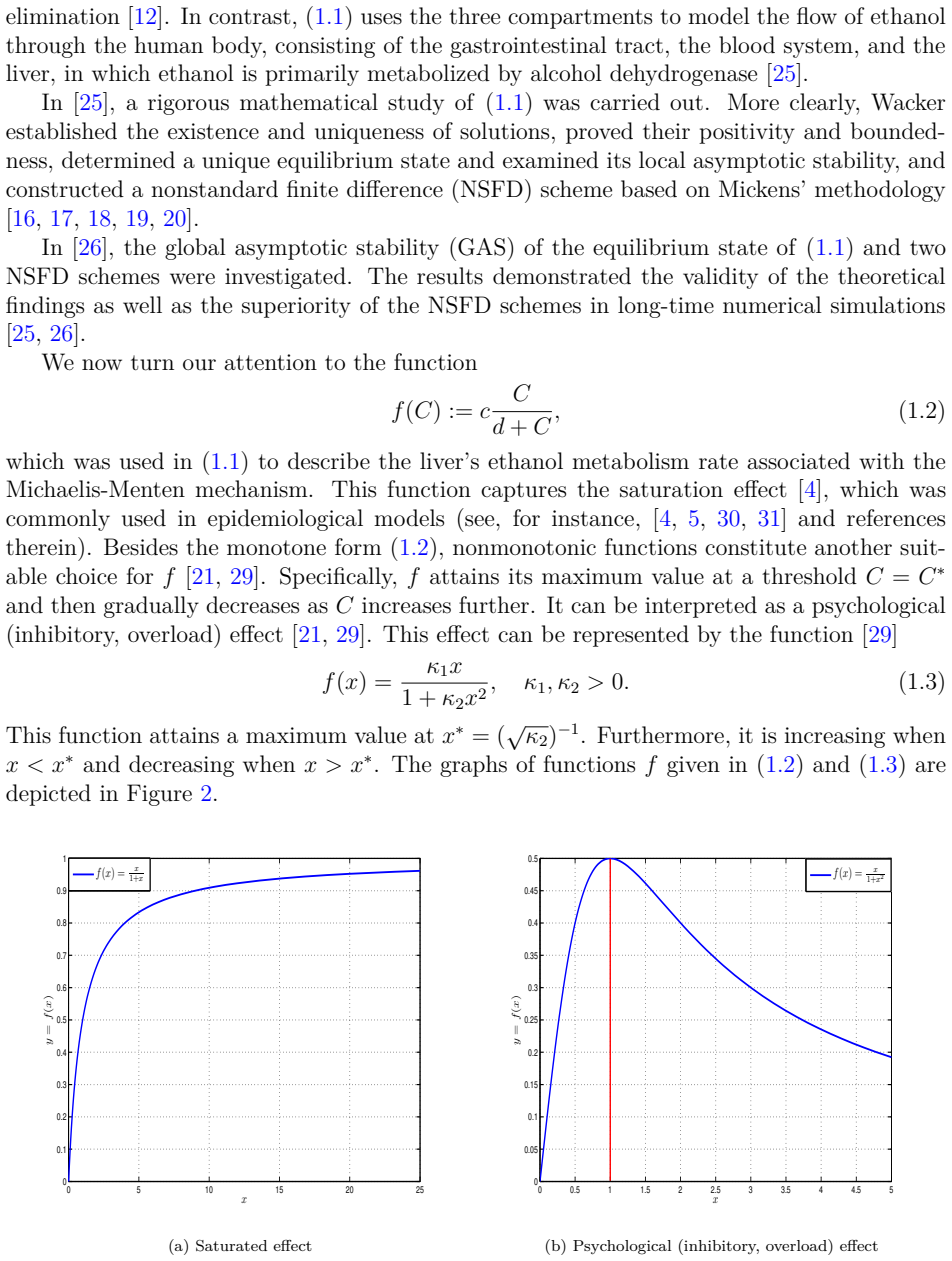
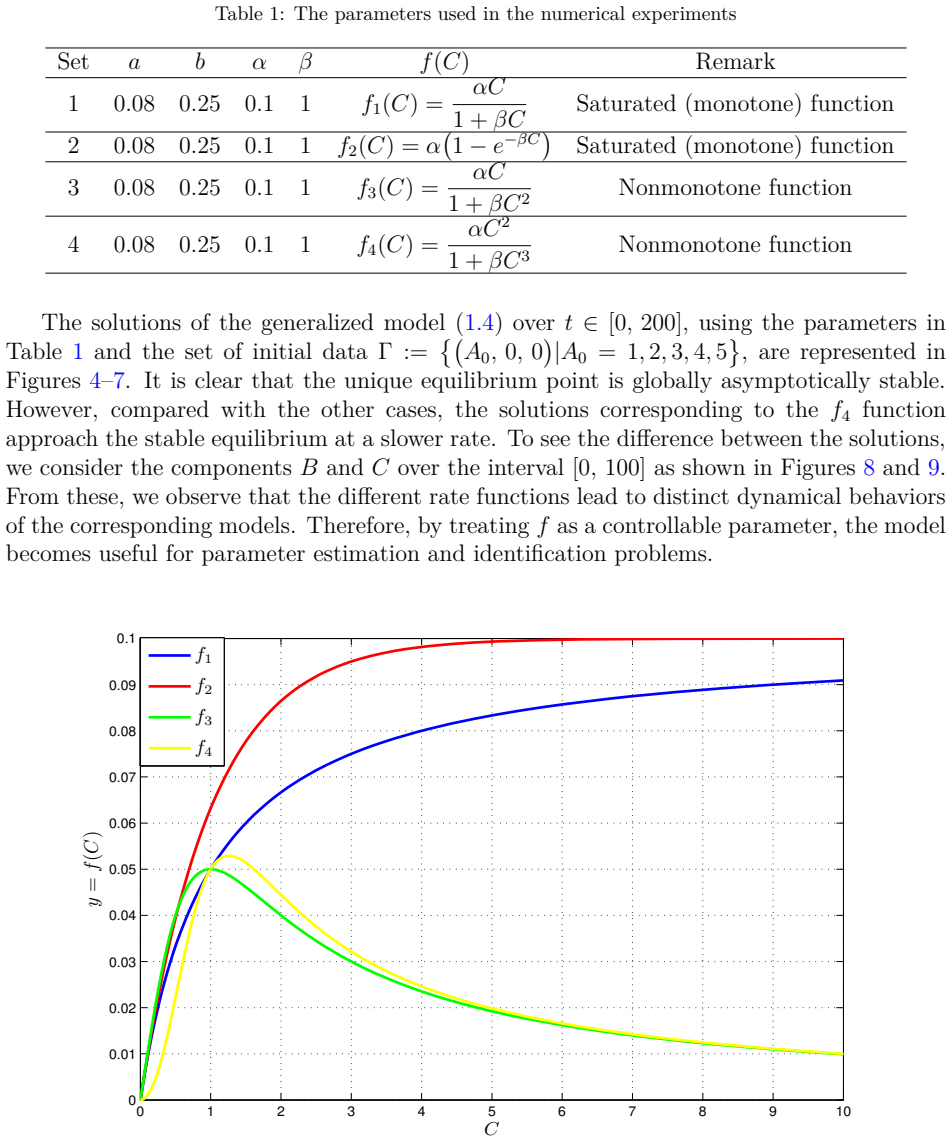
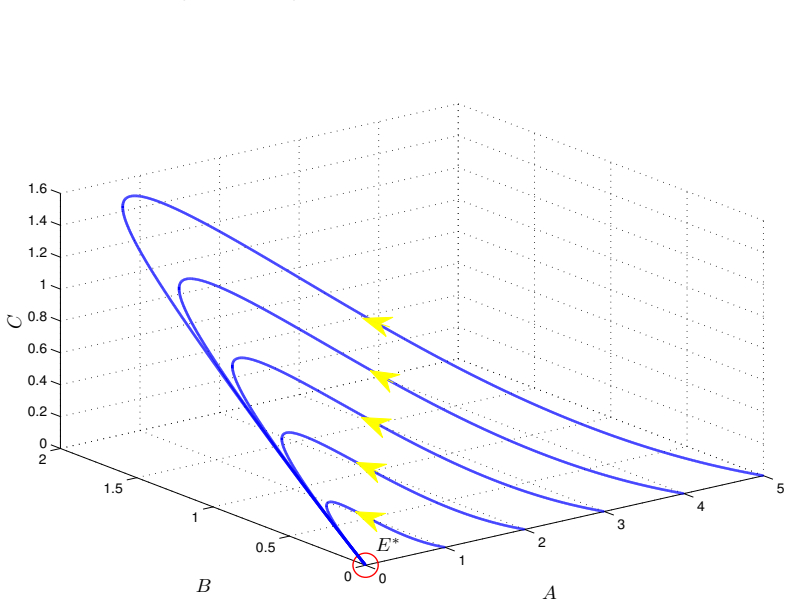
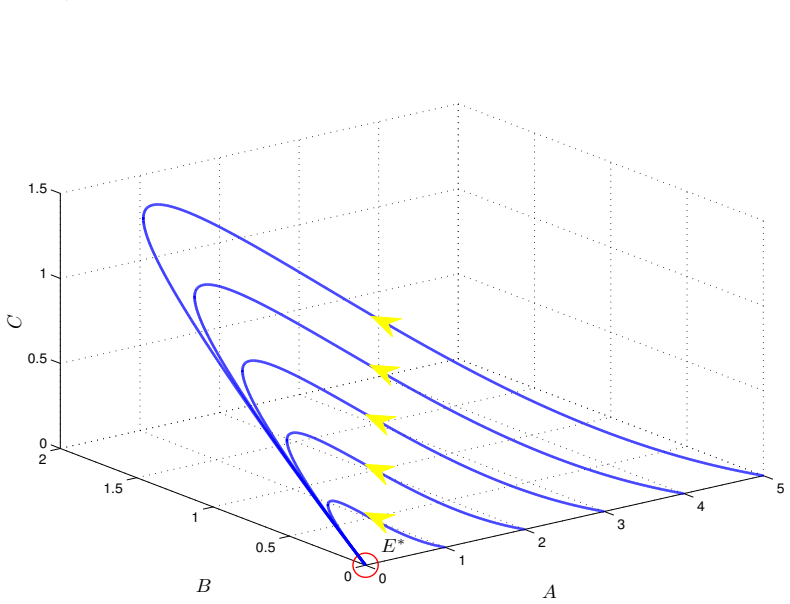
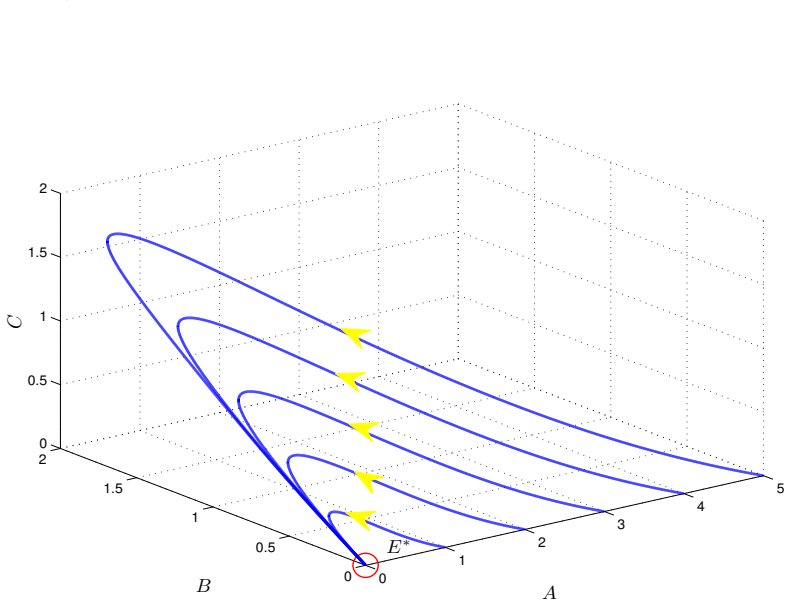
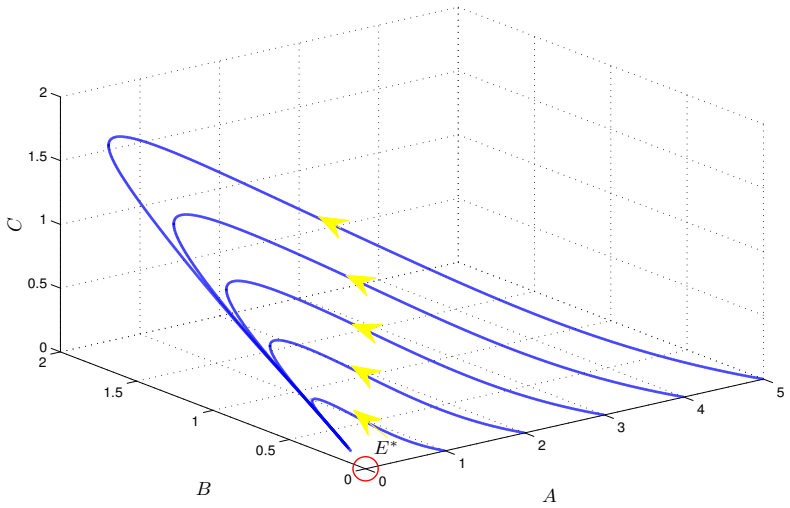
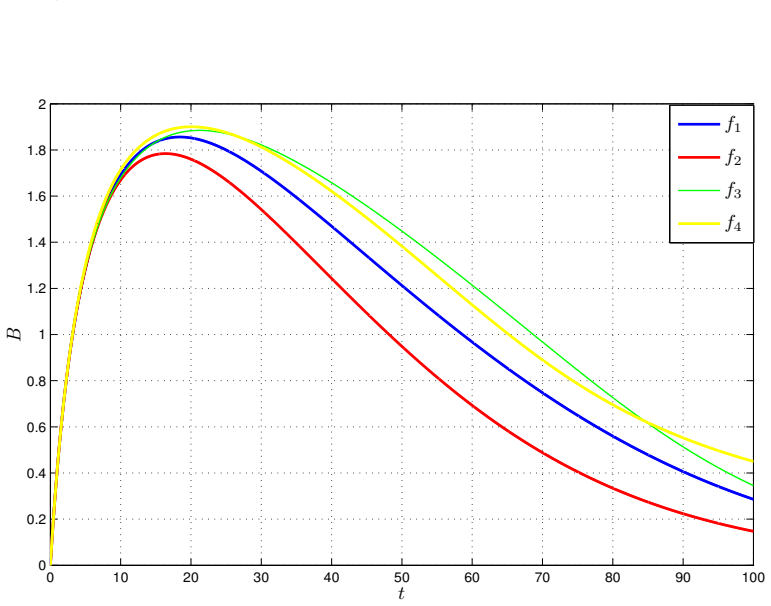
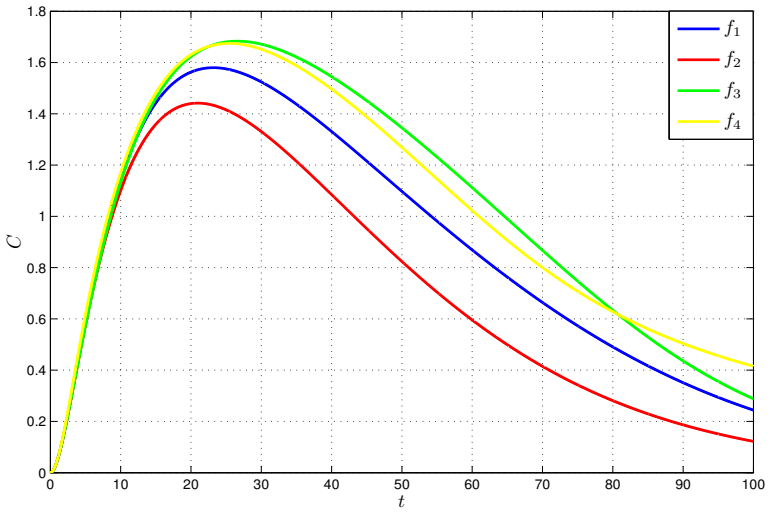
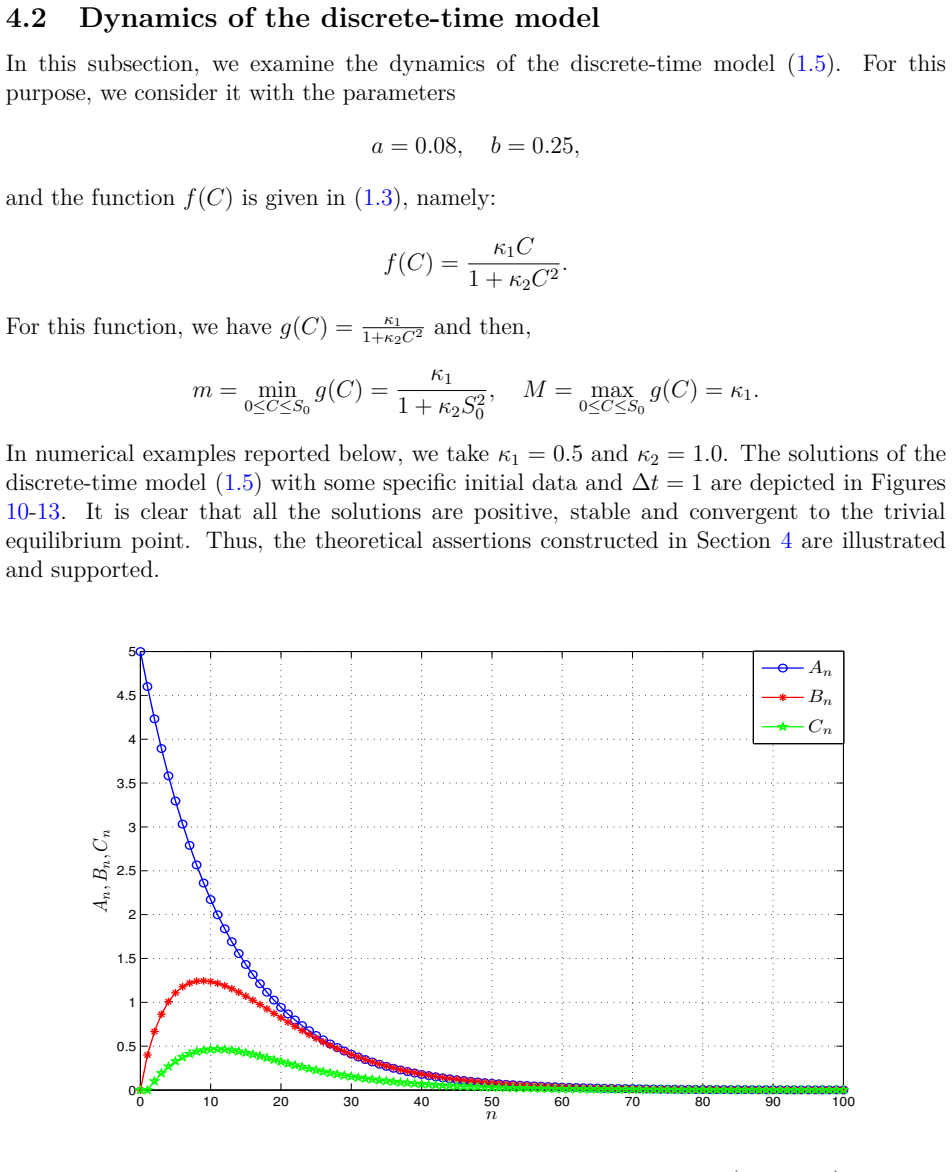
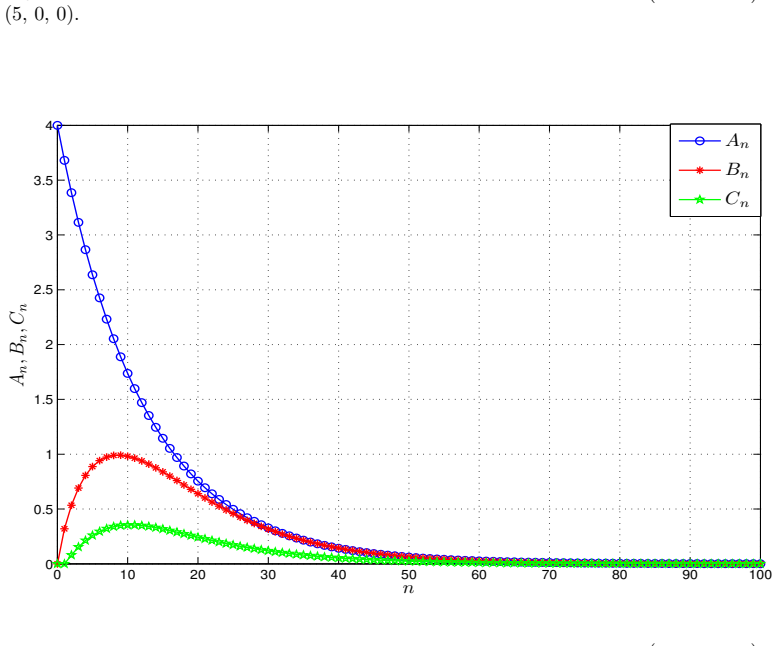
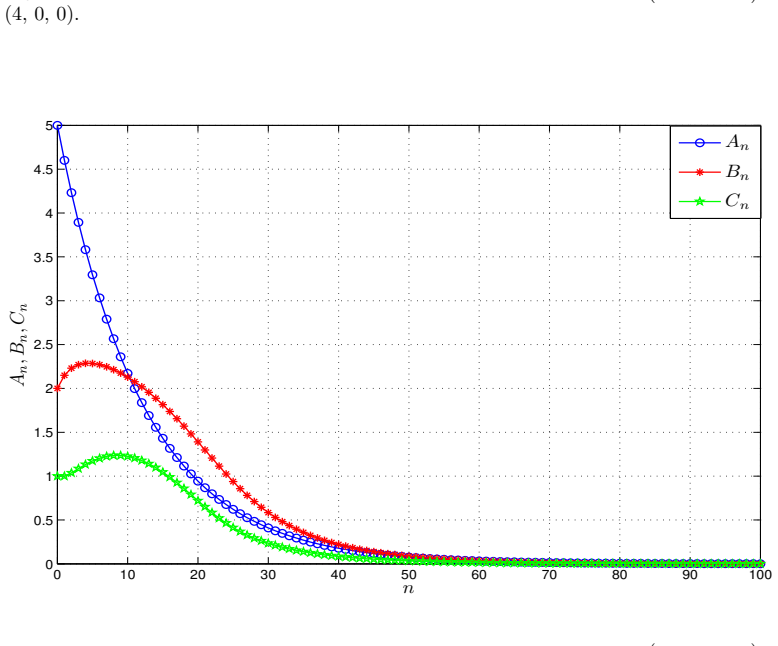
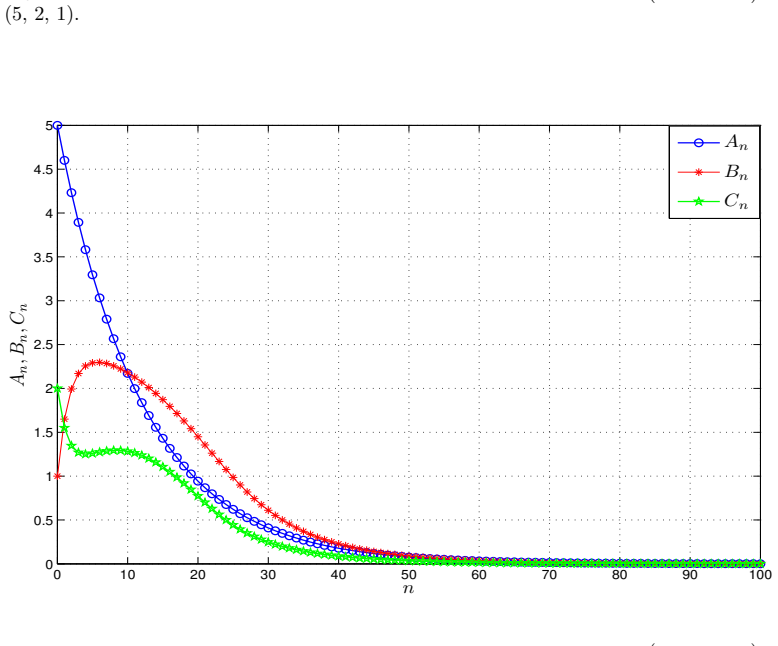
read the original abstract
We introduce a generalized continuous-time compartment model of ethanol metabolism in the human body that extends a recently developed framework. In the proposed model, we replace the Michaelis-Menten mechanism of the liver's ethanol metabolism rate with a general class of nonlinear rate functions. This modification provides greater modeling flexibility and enables the model to capture a wider range of hepatic ethanol metabolism dynamics. The qualitative behavior of the proposed ethanol metabolism model is analyzed rigorously. More specifically, we investigate the positivity and boundedness of solutions, as well as the global asymptotic stability (GAS) of the unique equilibrium point using an appropriate quadratic Lyapunov function. Second, we formulate a discrete-time counterpart of the proposed continuous-time model and investigate its dynamical properties. We show that, under an appropriate condition on the time step size, the discrete-time model faithfully reproduces the qualitative dynamical behavior of the corresponding continuous-time system. Lastly, we conduct a series of numerical experiments employing several ethanol metabolism rate functions to support the theoretical results.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper introduces a generalized continuous-time compartment model for ethanol metabolism, replacing the Michaelis-Menten liver rate with a general class of nonlinear rate functions to increase modeling flexibility. It claims to prove positivity and boundedness of solutions, existence of a unique equilibrium, and its global asymptotic stability via a quadratic Lyapunov function. A discrete-time counterpart is formulated, shown to reproduce the continuous dynamics under a suitable time-step restriction, and supported by numerical experiments with several rate functions.
Significance. If the technical hypotheses on the rate class are made explicit and the Lyapunov analysis is fully detailed and verified, the work would provide a flexible yet rigorously analyzed framework for compartment models in mathematical biology. The combination of continuous/discrete analysis and numerics is a positive feature, but the current presentation leaves the scope and verifiability of the central claims unclear.
major comments (3)
- [Model formulation] Model formulation section: The general class of nonlinear ethanol metabolism rate functions is introduced without an explicit list of the technical conditions (positivity, continuity, monotonicity or growth bounds) required for existence of a unique equilibrium and for the subsequent qualitative analysis. This premise is load-bearing for all claims of positivity, boundedness, and GAS.
- [Global asymptotic stability analysis] Global asymptotic stability section: The abstract and text refer to an 'appropriate quadratic Lyapunov function' but provide neither its explicit form nor the derivative calculation showing negative-definiteness along trajectories. Without this, the GAS claim cannot be verified.
- [Discrete-time model] Discrete-time model section: The 'appropriate condition on the time step size' that ensures the discrete system reproduces the continuous qualitative behavior is stated but not given explicitly or derived; this condition is central to the discrete-time fidelity claim.
minor comments (2)
- [Abstract] Abstract: The summary of results should reference the specific theorems or conditions rather than using vague phrases such as 'an appropriate condition on the time step size'.
- [Numerical experiments] Numerical experiments: The choice of parameter values and initial conditions in the simulations should be justified with reference to physiological ranges or prior literature.
Simulated Author's Rebuttal
We thank the referee for the careful reading and constructive comments, which help clarify the presentation of our results. We address each major comment below and will revise the manuscript to improve explicitness and verifiability while preserving the core contributions.
read point-by-point responses
-
Referee: [Model formulation] Model formulation section: The general class of nonlinear ethanol metabolism rate functions is introduced without an explicit list of the technical conditions (positivity, continuity, monotonicity or growth bounds) required for existence of a unique equilibrium and for the subsequent qualitative analysis. This premise is load-bearing for all claims of positivity, boundedness, and GAS.
Authors: We agree that the technical conditions on the rate functions should be stated explicitly rather than left implicit. In the revised manuscript we will add a dedicated paragraph or subsection listing the precise assumptions (positivity, continuity, monotonicity, and growth bounds) that are used to guarantee existence of a unique equilibrium and to carry out the positivity, boundedness, and stability analysis. revision: yes
-
Referee: [Global asymptotic stability analysis] Global asymptotic stability section: The abstract and text refer to an 'appropriate quadratic Lyapunov function' but provide neither its explicit form nor the derivative calculation showing negative-definiteness along trajectories. Without this, the GAS claim cannot be verified.
Authors: The referee correctly notes that the explicit quadratic Lyapunov function and the derivative calculation are not supplied in the current text. This omission prevents independent verification. In the revision we will state the precise form of the quadratic Lyapunov function and provide the full computation showing that its derivative is negative definite along trajectories under the model assumptions. revision: yes
-
Referee: [Discrete-time model] Discrete-time model section: The 'appropriate condition on the time step size' that ensures the discrete system reproduces the continuous qualitative behavior is stated but not given explicitly or derived; this condition is central to the discrete-time fidelity claim.
Authors: We acknowledge that the time-step restriction is mentioned only qualitatively. In the revised version we will derive and state the explicit upper bound on the time step h, together with the inequalities that guarantee preservation of positivity, boundedness, and global asymptotic stability for the discrete-time system. revision: yes
Circularity Check
No significant circularity
full rationale
The derivation consists of standard qualitative analysis (positivity, boundedness, GAS via quadratic Lyapunov) for a stated class of nonlinear rate functions satisfying explicit technical hypotheses (positivity, continuity, monotonicity/growth bounds). The discrete-time counterpart is shown to preserve the same properties under a time-step restriction. No step reduces by construction to a fitted parameter, self-definition, or load-bearing self-citation chain; the central claims remain conditional on the model-class assumptions and are independent of any internal renaming or ansatz smuggling. This is the normal case of a self-contained mathematical analysis.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption The ethanol metabolism rate functions are positive, continuous, and satisfy the growth and monotonicity conditions required for existence of a unique equilibrium and for the derivative of the quadratic Lyapunov function to be negative definite.
Reference graph
Works this paper leans on
-
[1]
L. J. S. Allen, Some discrete-time SI, SIR, and SIS epidemic models, Math. Biosci. 124 (1994) 83-125
1994
-
[2]
L. J. S. Allen, An introduction to mathematical biology, Pearson/Prentice Hall, 2007
2007
-
[3]
U. M. Ascher, L. R. Petzold, Computer Methods for Ordinary Differential Equations and Differential-Algebraic Equations, SIAM, Philadelphia, 1998
1998
-
[4]
Capasso, G
V. Capasso, G. Serio, A generalization of the Kermack-McKendrick deterministic epi- demic model, Mathematical Biosciences 42(1978) 43-61
1978
-
[5]
J. Cui, X. Mu, H. Wan, Saturation recovery leads to multiple endemic equilibria and backward bifurcation, Journal of Theoretical Biology 254 (2008) 275-283. 17
2008
-
[6]
J.H.P. Dawes, M.O. Souza, A derivation of Holling’s type I, II and III functional responses in predator-prey systems, Journal of Theoretical Biology 327:11–22, 2013, https://doi.org/10.1016/j.jtbi.2013.02.017
-
[7]
H.J. Edenberg, J.N. McClintick, Alcohol Dehydrogenases, Aldehyde Dehydrogenases, and Alcohol Use Disorders: A Critical Review, Alcoholism: Clinical and Experimental Research 42(12):2281–2297, 2018, https://doi.org/10.1111/acer.13904
-
[8]
Elaydi, An introduction to difference equations, Springer-Verlag, New York, 2005
S. Elaydi, An introduction to difference equations, Springer-Verlag, New York, 2005
2005
-
[9]
G. Kalinkat, B.C. Rall, S.F. Uiterwall, W. Uszko, Empirical evidence of type III func- tional responses and why it remains rare, Frontiers in Ecology and Evolution 11:1033818, 2023, https://doi.org/10.3389/fevo.2023.1033818
-
[10]
H. K. Khalil, Nonlinear systems, Third Edition, Prentice Hall, 2002
2002
-
[11]
L. M. Ladino, C. Mammana, E. Michetti, J. C. Valverde, Discrete time population dynamics of a two-stage species with recruitment and capture, Chaos, Solitons and Fractals 85 (2016) 143-150
2016
-
[12]
M. D. Levitt, D. G. Levitt, Use of a Two-Compartmental Model to Assess Pharma- cokinetics of Human Ethanol Metabolism, Alcoholism: Clinical and Experimental Re- search 22, no. 8 (1998): 1680-1688,https://doi.org/10.1111/j.1530-0277.1998. tb03966.x
-
[13]
J. Li, Z. Ma , F. Brauer, Global analysis of discrete-time SI and SIS epidemic models, Mathematical Biosciences and Engineering 2007(4) 699-710
2007
-
[14]
Ludwin, Blood Alcohol Content, Undergraduate Journal of Mathematical Modeling: One + two 3, no
C. Ludwin, Blood Alcohol Content, Undergraduate Journal of Mathematical Modeling: One + two 3, no. 2 (2011): 1, https://doi.org/10.5038/2326-3652.3.2.1
-
[15]
R. M. May, Simple mathematical models with very complicated dynamics, Nature 261(1976) 459-467
1976
-
[16]
R. E. Mickens, Nonstandard Finite Difference Models of Differential Equations, World Scientific, Singapore, 1994
1994
-
[17]
R. E. Mickens, Applications of Nonstandard Finite Difference Schemes, World Scientific, Singapore, 2000
2000
-
[18]
R. E. Mickens, Dynamic consistency: a fundamental principle for constructing nonstan- dard finite difference schemes for differential equations, Journal of Difference Equations and Applications 11 (2005) 645-653
2005
-
[19]
R. E. Mickens, Advances in the Applications of Nonstandard Finite Difference Schemes, World Scientific, Singapore, 2005
2005
-
[20]
R. E. Mickens, Nonstandard Finite Difference Schemes: Methodology and Applications, World Scientific, 2020. 18
2020
-
[21]
Ruana, W
S. Ruana, W. Wang, Dynamical behavior of an epidemic model with a nonlinear inci- dence rate, Journal of Differential Equations 188(2003) 135-163
2003
-
[22]
Seibert, R
P. Seibert, R. Suarez, Global stabilization of nonlinear cascade systems, Syst. Contr. Lett. 14 (1990), 347–352
1990
-
[23]
H. L. Smith and P. Waltman, The Theory of the Chemostat: Dynamics of Microbial Competition, Cambridge University Press, 2009
2009
-
[24]
A. M. Stuart, A. R. Humphries, Dynamical systems and numerical analysis, Cambridge university press, 1998
1998
-
[25]
B. Wacker, Analysis of a Finite-Difference Method Based on Nonlocal Approximations for a Nonlinear, Extended Three-Compartmental Model of Ethanol Metabolism in the Human Body, Mathematical Methods in the Applied Sciences 48 (2025), 9975-9992
2025
-
[26]
Wacker, Global asymptotic stability of one time-continuous and two time-discrete models for ethanol metabolism in the human body, Aequat
B. Wacker, Global asymptotic stability of one time-continuous and two time-discrete models for ethanol metabolism in the human body, Aequat. Math. 99 (2025), 2401- 2422
2025
-
[27]
X. Wang, Z. Wang, H. Shen, Dynamical analysis of a discrete-time SIS epidemic model on complex networks, Applied Mathematics Letters 94 (2019) 292-299
2019
-
[28]
X. Wu, Z. Zhou, F. Xie, Multi-scale dynamics of a piecewise-smooth Bazykin’s prey- predator system, Nonlinear Dynamics 113:1969–1981, 2025, https://doi.org/10. 1007/s11071-024-10292-9
1969
-
[29]
D. Xiao, S. Ruan, Global analysis of an epidemic model with nonmonotone incidence rate, Mathematical Biosciences 208 (2007) 419-429
2007
-
[30]
R. Xu, Z. Ma, Global stability of a delayed SEIRS epidemic model with saturation incidence rate, Nonlinear Dyn (2010) 61: 229-239
2010
-
[31]
Zhang, X
X. Zhang, X. Liu, Backward bifurcation of an epidemic model with saturated treatment function, J. Math. Anal. Appl. 348 (2008) 433-443. 19
2008
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.