Cell-scale degradation of peritumoural extracellular matrix fibre network and its role within tissue-scale cancer invasion
Pith reviewed 2026-05-25 10:36 UTC · model grok-4.3
The pith
Modeling ECM fibre degradation at both tumour bulk and cell-scale edge shows the MT1-MMP/MMP-2 cascade affects tissue-scale invasion.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
The paper proposes a new multiscale modelling framework in which the degradation of peritumoural extracellular matrix fibres occurs both at macro-scale within the tumour bulk and explicitly at micro-scale in the neighbourhood of the tumour boundary due to the MT1-MMP/MMP-2 cascade. This framework couples the micro-dynamics of matrix degrading enzyme fluxes and fibre degradation to the continuous rearrangement of naturally oriented ECM fibres to investigate the overall effect on tumour progression.
What carries the argument
The multiscale framework that relates micro-scale MDE proteolytic dynamics at the tumour interface to macro-scale fibre rearrangement in the tumour bulk.
If this is right
- Local degradation at the invasive edge creates space for cancer cells to progress.
- The MT1-MMP/MMP-2 interactions with ECM fibres affect the overall tumour invasion pattern.
- Coupling cell-scale and tissue-scale processes leads to more accurate predictions of invasion outcomes.
- The model highlights the role of fibre orientation rearrangement in conjunction with enzymatic degradation.
Where Pith is reading between the lines
- Therapies aimed at boundary enzymes might produce distinct invasion control compared to bulk targeting.
- The model could be tested against measurements of local fibre degradation rates in cell culture at the tumour-ECM interface.
- Extension to three-dimensional geometries might uncover additional directional effects on spread.
Load-bearing premise
The micro-dynamics of MDE fluxes and fibre degradation in the cell-scale neighbourhood of the tumour boundary can be coupled to the continuous rearrangement of naturally oriented ECM fibres in the tumour bulk to determine tissue-scale invasion outcomes.
What would settle it
An experiment or simulation in which micro-scale fibre degradation at the tumour edge is blocked while bulk degradation continues, yet invasion patterns and speed remain unchanged from the coupled case.
Figures





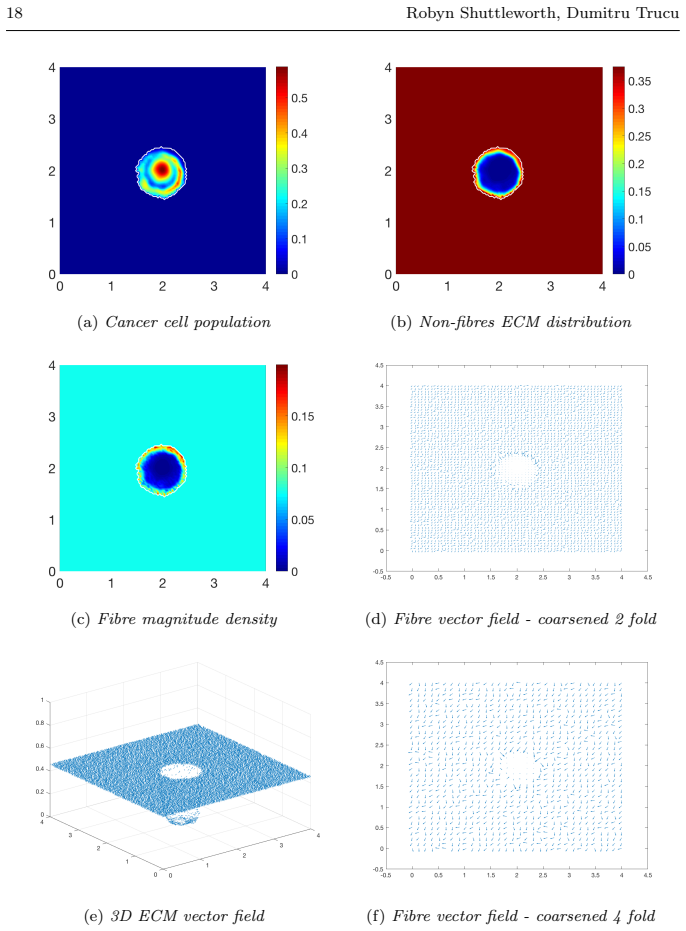


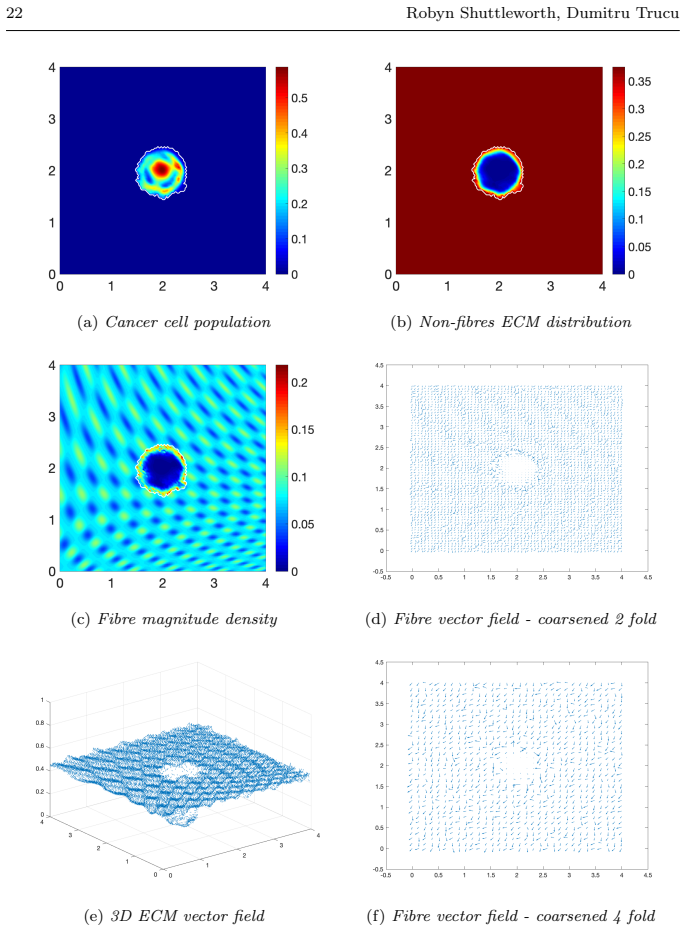



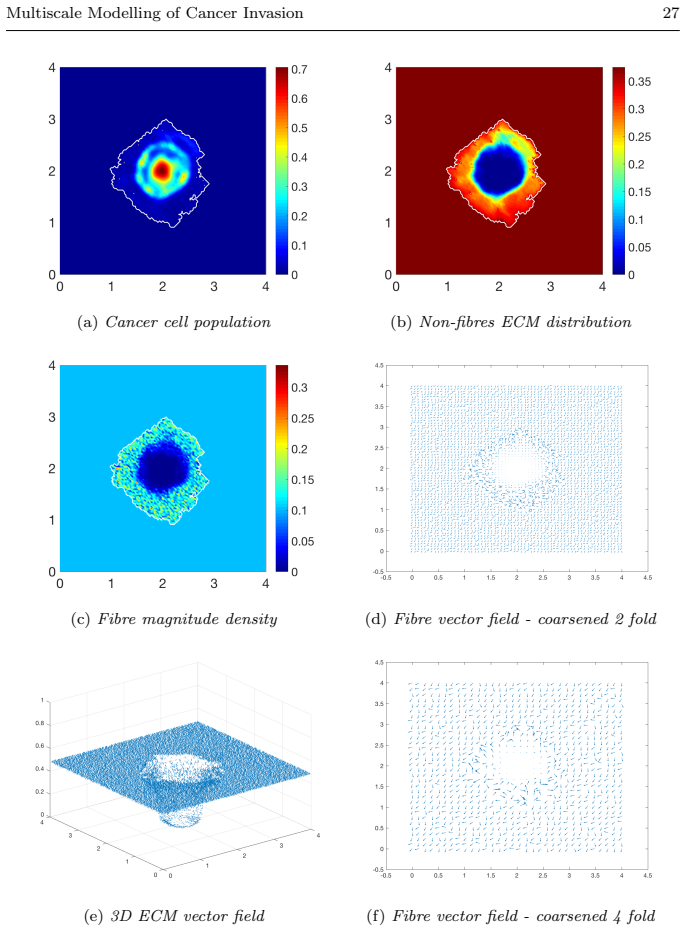
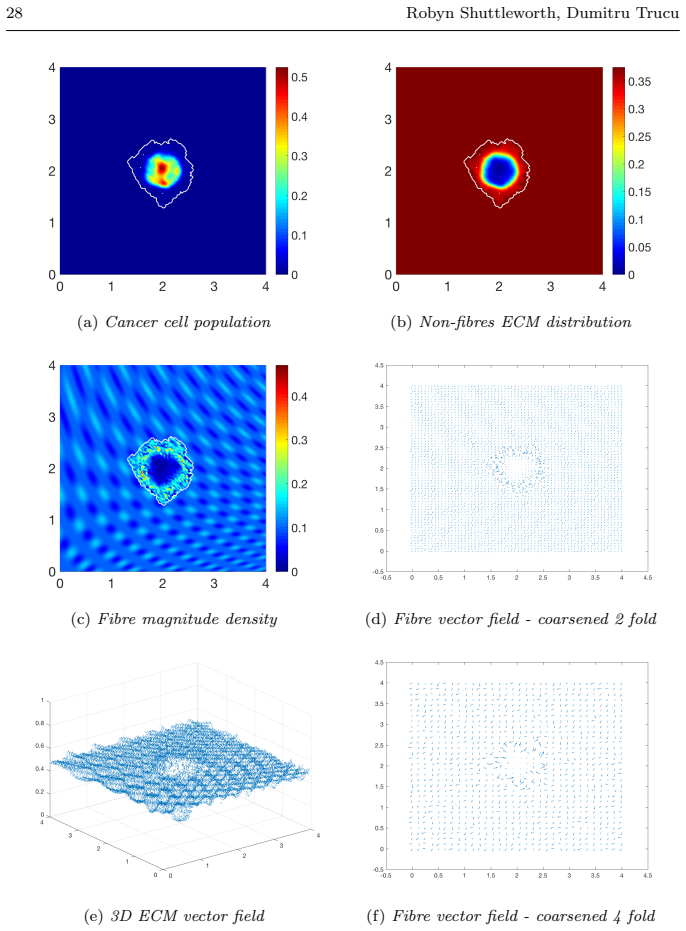




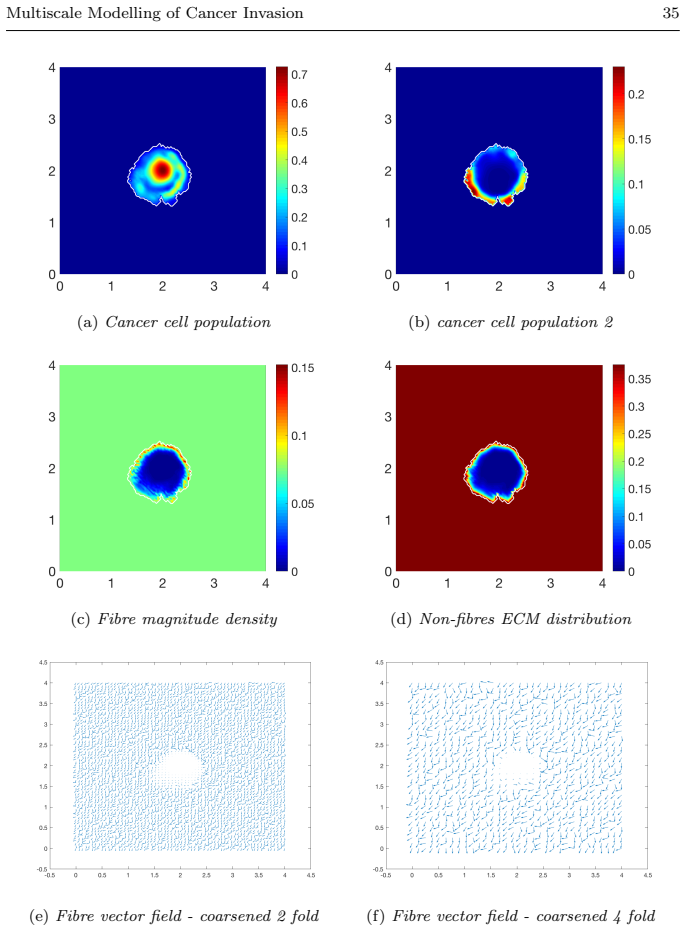
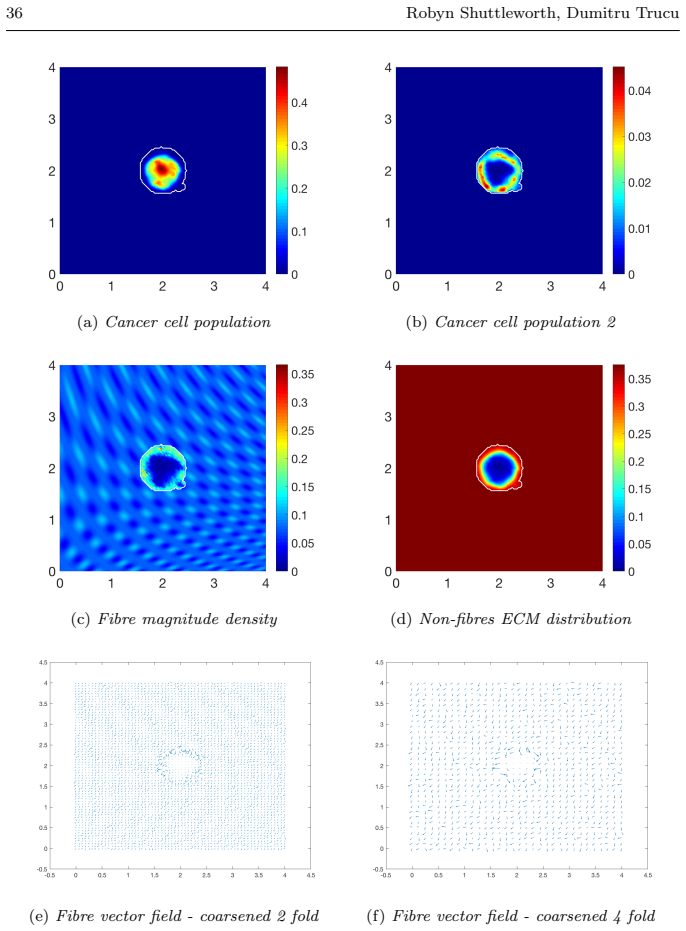

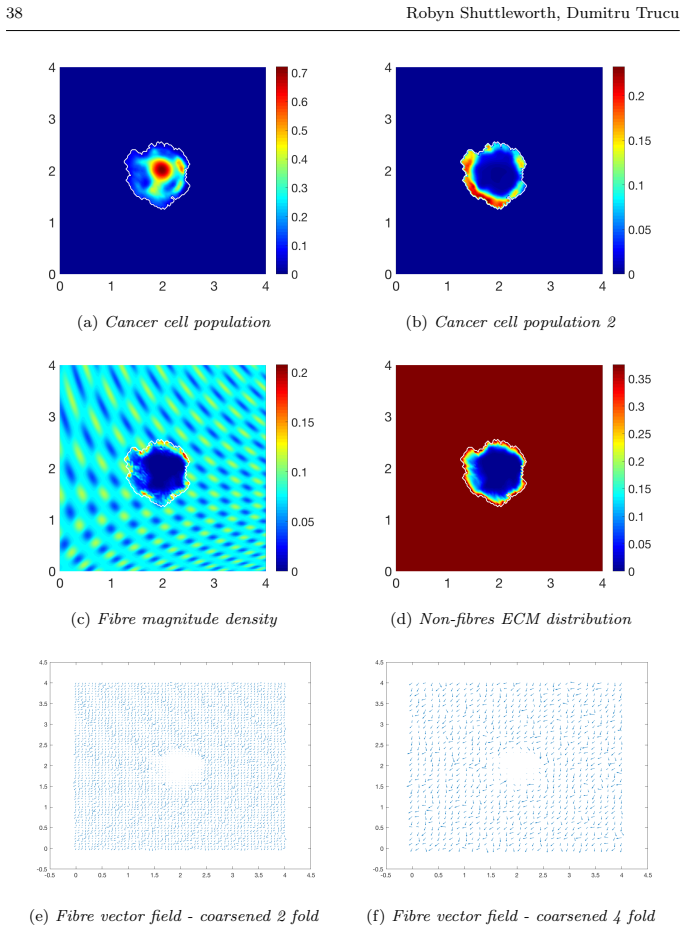
read the original abstract
Local cancer invasion of tissue is a complex, multiscale process which plays an essential role in tumour progression. Occurring over many different temporal and spatial scales, the first stage of invasion is the secretion of matrix degrading enzymes (MDEs) by the cancer cells that consequently degrade the surrounding extracellular matrix (ECM). This process is vital for creating space in which the cancer cells can progress and it is driven by the activities of specific matrix metalloproteinases (MMPs). In this paper, we consider the key role of two MMPs by developing further the novel two-part multiscale model introduced in [33] to better relate at micro-scale the two micro-scale activities that were considered there, namely, the micro-dynamics concerning the continuous rearrangement of the naturally oriented ECM fibres within the bulk of the tumour and MDEs proteolytic micro-dynamics that take place in an appropriate cell-scale neighbourhood of the tumour boundary. Focussing primarily on the activities of the membrane-tethered MT1-MMP and the soluble MMP-2 with the fibrous ECM phase, in this work we investigate the MT1-MMP/MMP-2 cascade and its overall effect on tumour progression. To that end, we will propose a new multiscale modelling framework by considering the degradation of the ECM fibres not only to take place at macro-scale in the bulk of the tumour but also explicitly in the micro-scale neighbourhood of the tumour interface as a consequence of the interactions with molecular fluxes of MDEs that exercise their spatial dynamics at the invasive edge of the tumour.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript extends the two-part multiscale model from [33] by incorporating the MT1-MMP/MMP-2 cascade and its interactions with the fibrous ECM phase. It proposes a new framework in which ECM fibre degradation occurs both at macro-scale within the tumour bulk (via continuous fibre rearrangement) and explicitly at micro-scale in a cell-scale neighbourhood of the tumour interface (via MDE fluxes), with the goal of determining the cascade's overall effect on tissue-scale invasion outcomes.
Significance. If the scale-coupling is shown to transmit proteolytic activity without introducing artifacts, the work would strengthen the link between specific MMP molecular dynamics and observable invasion patterns, building directly on the prior model in [33].
major comments (1)
- Abstract (paragraph on the new multiscale modelling framework): the claim that micro-scale MDE fluxes and fibre degradation can be coupled to macro-scale fibre rearrangement to determine invasion outcomes is load-bearing, yet the provided text supplies no equations, parameter values, or simulation outputs, preventing verification that the cascade effect crosses scales without loss or post-hoc adjustment.
Simulated Author's Rebuttal
We thank the referee for their review and for highlighting the importance of verifying the scale-coupling in our proposed multiscale framework. We address the single major comment below.
read point-by-point responses
-
Referee: [—] Abstract (paragraph on the new multiscale modelling framework): the claim that micro-scale MDE fluxes and fibre degradation can be coupled to macro-scale fibre rearrangement to determine invasion outcomes is load-bearing, yet the provided text supplies no equations, parameter values, or simulation outputs, preventing verification that the cascade effect crosses scales without loss or post-hoc adjustment.
Authors: The abstract is intended as a concise summary of the modelling approach and its motivation. The full manuscript develops the coupling explicitly: Section 2 recalls the two-part framework from [33] and introduces the MT1-MMP/MMP-2 cascade; Section 3 derives the micro-scale MDE flux equations and the resulting fibre degradation term at the tumour interface; Section 4 presents the macro-scale fibre rearrangement equations together with the transmission conditions that link the two scales; and Section 5 reports numerical simulations with the specific parameter sets used, showing invasion outcomes that arise directly from the coupled system without additional tuning. These sections contain the governing PDEs, boundary conditions, parameter tables, and simulation outputs that allow verification of scale transmission. revision: no
Circularity Check
No significant circularity detected
full rationale
The paper proposes a new multiscale modelling framework that extends the two-part model from reference [33] by the same authors, focusing on coupling micro-scale MT1-MMP/MMP-2 driven fibre degradation at the tumour interface with macro-scale ECM rearrangement. No equations or derivations are presented in the provided material that reduce predictions to fitted inputs by construction, self-definitions, or load-bearing self-citations whose validity depends on the current work. The central claim concerns the overall effect of the cascade across scales, which is introduced as an independent modelling contribution rather than a renaming or forced outcome of prior assumptions. The derivation chain is therefore self-contained against external benchmarks.
Axiom & Free-Parameter Ledger
free parameters (2)
- MDE secretion and diffusion rates
- fibre degradation rate constants for MT1-MMP/MMP-2
axioms (2)
- domain assumption ECM fibres possess a natural orientation that undergoes continuous rearrangement inside the tumour bulk
- domain assumption MDEs exercise spatial dynamics that can be localised to a cell-scale neighbourhood of the tumour boundary
Reference graph
Works this paper leans on
-
[1]
Andasari, V., Gerisch, A., Lolas, G., South, A., Chaplain, M.A.J.: Mathematical mod- eling of cancer cell invasion of tissue: biological insight from mathematical analy- sis and computational simulation. J. Math. Biol. 63(1), 141–171 (2011). DOI 10.1007/s00285-010-0369-1
-
[2]
Anderson, A.R.A.: A hybrid mathematical model of solid tumour invasion: the im- portance of cell adhesion. Math. Medic. Biol. 22(2), 163–186 (2005). DOI 10.1093/ imammb/dqi005
work page 2005
-
[3]
Anderson, A.R.A., Chaplain, M.A.J., Newman, E.L., Steele, R.J.C., Thompson, A.M.: Mathematical modelling of tumour invasion and metastasis. J. Theor. Med. 2(2), 129– 154 (2000). DOI 10.1080/10273660008833042
-
[4]
Anderson, A.R.A., Rejniak, K.A., Gerlee, P., Quaranta, V.: Modelling of cancer growth, evolution and invasion: Bridging scales and models. Math. Model. Nat. Phenom. 2(3), 1–29 (2007)
work page 2007
-
[5]
Bhagavathula, N., Hanosh, A.W., Nerusu, K.C., Appelman, H., Chakrabarty, S., Varani, J.: Regulation of e-cadherin and β-catenin by Ca 2+ in colon carcinoma is dependent on calcium-sensing receptor expression and function. Int. J. Cancer 121, 1455–1462 (2007). DOI 10.1002/ijc.22858
-
[6]
Bitsouni, V., Chaplain, M.A.J., Eftimie, R.: Mathematical modelling of cancer invasion: The multiple roles of tgf- β pathway on tumour proliferation and cell adhesion. Math. Model. Meth. Appl. Sci. 27(10), 1929–1962 (2017)
work page 1929
-
[7]
Cavallaro, U., Christofori, G.: Cell adhesion in tumor invasion and metastasis: loss of the glue is not enough. Biochim. Biophys. Acta. 1552(1), 39–45 (2001). DOI 10.1016/S0304-419X(01)00038-5
-
[8]
Math Mod Meth Appl S 21, 719–743 (2011)
Chaplain, M.A.J., Lachowicz, M., Szyma´ nska, Z., Wrzosek, D.: Mathematical modelling of cancer invasion: The importance of cell-cell adhesion and cell-matrix adhesion. Math Mod Meth Appl S 21, 719–743 (2011). DOI 10.1142/S0218202511005192
-
[9]
Chaplain, M.A.J., Lolas, G.: Mathematical modelling of cancer cell invasion of tissue: The role of the urokinase plasminogen activation system. Math. Model. Meth. Appl. Sci. 15, 1685–1734 (2005). DOI 10.1142/S0218202505000947
-
[10]
Chaplain, M.A.J., Lolas, G.: Mathematical modelling of cancer invasion of tissue: Dy- namic heterogeneity. Netw. Heterog. Media. 1(3), 399–439 (2006). DOI 10.3934/nhm. 2006.1.399
work page doi:10.3934/nhm 2006
-
[11]
Chaplain, M.A.J., McDougall, S.R., Anderson, A.R.A.: Mathematical modelling of tumor-induced angiogenesis. Annu. Rev. Biomed. Eng. 8, 233–257 (2006). DOI 10.1146/annurev.bioeng.8.061505.095807
-
[12]
Chauviere, A., Hillen, T., Preziosi, L.: Modeling cell movement in anisotropic and het- erogeneous network tissues. Netw. Heterog. Media. 2(2), 333–357 (2007)
work page 2007
-
[13]
Deakin, N.E., Chaplain, M.A.J.: Mathematical modelling of cancer cell invasion: the role of membrane-bound matrix metalloproteinases. Front. Oncol. 3(70), 1–9 (2013)
work page 2013
-
[14]
Domschke, P., Trucu, D., Gerisch, A., Chaplain, M.A.J.: Mathematical modelling of cancer invasion: Implications of cell adhesion variability for tumour infiltrative growth patterns. J. Theor. Biol. 361, 41–60 (2014)
work page 2014
-
[15]
Gerisch, A., Chaplain, M.A.J.: Mathematical modelling of cancer cell invasion of tissue: Local and non-local models and the effect of adhesion. J. Theor. Biol. 250(4), 684–704 (2008). DOI 10.1016/j.jtbi.2007.10.026
-
[16]
Gu, Z., Liu, F., Tonkova, E.A., Lee, S.Y., Tschumperlin, D.J., Brenner, M.B., Ginsberg, M.H.: Soft matrix is a natural stimulator for cellular invasiveness. Mol. Biol. Cell.25(4), 457–469 (2014). DOI 10.1091/mbc.e13-05-0260
-
[17]
Hillen, T.: M5 mesoscopic and macroscopic models for mesenchymal motion. J. Math. Biol. 53(4), 585–616 (2006). DOI 10.1007/s00285-006-0017-y
-
[18]
Hofer, A.M., Curci, S., Doble, M.A., Brown, E.M., Soybel, D.I.: Intercellular commu- nication mediated by the extracellular calcium-sensing receptor. Nat. Cell. Biol. 2(7), 392–398 (2000). DOI 10.1038/35017020
-
[19]
Prentice-Hall (1987) 44 Robyn Shuttleworth, Dumitru Trucu
Hughes, T.J.R.: The Finite Element Method: Linear static and dynamic finite element analysis. Prentice-Hall (1987) 44 Robyn Shuttleworth, Dumitru Trucu
work page 1987
-
[20]
Humphries, J.D., Byron, A., Humphries, M.J.: Integrin ligands at a glance. J. Cell. Sci. 119, 3901–3903 (2006). DOI 10.1242/jcs.03098
-
[21]
Ko, K.S., Arora, P.D., Bhide, V., Chen, A., McCulloch, C.A.G.: Cell-cell adhesion in human fibroblasts requires calcium signalling. J. Cell. Sci. 114(6), 1155–1167 (2001)
work page 2001
-
[22]
Lodish, H., Berk, A., Zipursky, L., J.E.Darnell, Matsudaira, P., Baltimore, D.: Molecular Cell Biology, 4 edn. New York: W.H. Freeman (2000)
work page 2000
-
[23]
Painter, K.J.: Modelling cell migration strategies in the extracellular matrix. J. Math. Biol. 58(4), 511 (2008). DOI 10.1007/s00285-008-0217-8
-
[24]
Peng, L., Trucu, D., Lin, P., Thompson, A., Chaplain, M.A.J.: A multiscale mathemat- ical model of tumour invasive growth. Bull. Math. Biol. 79(3), 389–429 (2017)
work page 2017
-
[25]
Perumpanani, A.J., Simmons, D.L., Gearing, A.J.H., Miller, K.M., Ward, G., Norbury, J., Schneemann, M., Sherratt, J.A.: Extracellular matrix-mediated chemotaxis can im- pede cell migration. Proc. Biol. Sci. 265(1413), 2347–2352 (1998)
work page 1998
-
[26]
Provenzano, P.P., Eliceiri, K.W., Campbell, J.M., Inman, D.R., White, J.G., Keely, P.J.: Collagen reorganization at the tumor-stromal interface facilitates local invasion. BMC Med. 4(38) (2006). DOI 10.1186/1741-7015-4-38
-
[27]
Provenzano, P.P., Inman, D.R., Eliceiri, K.W., Knittel, J.G., Yan, L., Rueden, C.T., White, J.G., Keely, P.J.: Collagen density promotes mammary tumour initiation and progression. BMC Med. 6(11) (2008). DOI 10.1186/1741-7015-6-11
-
[28]
Ramis-Conde, I., Chaplain, M.A.J., Anderson, A.R.A.: Mathematical modelling of tu- mour invasion and metastasis. Math. Comput. Model. 47(5-6), 533–545 (2008)
work page 2008
-
[29]
Ruangpanit, N., Chan, D., Holmbeck, K., Birkedal-Hansen, H., Polarek, J., Yang, C., Bateman, J.F., Thompson, E.W.: Gelatinase a (mmp-2) activation by skin fibroblasts: dependence on mt1-mmp expression and fibrillar collagen form. Matrix Biol. 20(3), 193–203 (2001). DOI 10.1016/S0945-053X(01)00135-4
-
[30]
Seiki, M., Yana, I.: Roles of pericellular proteolysis by membrane type-1 matrix metal- loproteinase in cancer invasion and angiogenesis. Cancer Metast. Rev. 94(7), 569–574 (2003)
work page 2003
-
[31]
Shuttleworth, R., Trucu, D.: Cell Movement Modelling and Applications, chap. Two- scale Moving Boundary Dynamics of Cancer Invasion: Heterotypic Cell Popula- tions Evolution in Heterogeneous ECM, pp. 1–26. Springer (2018). DOI 10.1007/ 978-3-319-96842-1
work page 2018
-
[32]
Shuttleworth, R., Trucu, D.: Multiscale dynamics of a hetertypic cancer cell population within a fibrous extracellular matrix (2019)
work page 2019
-
[33]
Shuttleworth, R., Trucu, D.: Multiscale modelling of fibres dynamics and cell adhesion within moving boundary cancer invasion. Bull. Math. Biol. (2019). DOI 10.1007/ s11538-019-00598-w
work page 2019
-
[34]
Szyma´ nska, Z., Morales-Rodrigo, C., Lachowicz, M., Chaplain, M.A.J.: Mathematical modelling of cancer invasion of tissue: the role and effect of nonlocal interactions. Math. Model. Meth. Appl. Sci. 19(2), 257–281 (2009)
work page 2009
-
[35]
Tam, E.M., Moore, T.R., Butler, G.S., Overall, C.M.: Characterization of the distint collagen binding, helicase and cleavage mechanisms of matrix metalloproteinases 2 and 14 (gelatinase a and mt1-mmp). J. Biol. Chem. 279(41), 43,336–43,344 (2004)
work page 2004
-
[36]
Thomson, E.W., Yu, M., Bueno, J., Jin, L., Maiti, S.N., Palao-Marco, F.L., Pulyaeva, H., Tamborlane, J.W., Tirgari, R., et al., I.W.: Collagen induced mmp-2 activation in human breast cancer. Breast Cancer Res. Treat. 31(2-3), 357–370 (1994)
work page 1994
-
[37]
Trucu, D., Lin, P., Chaplain, M.A.J., Wang, Y.: A multiscale moving boundary model arising in cancer invasion. Multiscale Model. Simul. 11(1), 309–335 (2013)
work page 2013
-
[38]
Van Doren, S.R.: Matrix metallopreinase interactions with collagen and elastin. Matrix Biol. 44-46, 224–231 (2015). DOI 10.1016/j.matbio.2015.01.005
-
[39]
Wijnhoven, B.J., Dinjens, W.N., Pignatelli, M.: E-cadherin-catenin cell-cell adhesion complex and human cancer. Brit. J. Surg. 87(8), 992–1005 (2000)
work page 2000
-
[40]
Wolf, K., te Lindert, M., Krause, M., Alexander, S., te Riet, J., Willis, A.L., Hoffman, R.M., Figdor, C.G., Weiss, S.J., Friedl, P.: Physical limits of cell migration: Control by ECM space and nuclear deformation and tuning by proteolysis and traction force. J. Cell. Biol. 201(7), 1069–1084 (2013). DOI 10.1083/jcb.201210152
-
[41]
Yosida, K.: Functional Analysis, 6th edn. Springer-Verlag (1980)
work page 1980
-
[42]
Zigrino, P., Drescher, C., Mauch, C.: Collagen-induced proMMP-2 activation by MT1- MMP in human dermal fibroblasts and the possible role of α2β1 integrins. Euro. J. Cell. Biol. 80, 68–77 (2001)
work page 2001
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.