Global dynamics and spatio-temporal patterns of predator-prey systems with density-dependent motion
Pith reviewed 2026-05-25 09:20 UTC · model grok-4.3
The pith
Predator-prey systems with density-dependent preytaxis have globally bounded classical solutions whose homogeneous equilibria are globally stable under parameter conditions.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
For the predator-prey system with prey-density-dependent motility function, classical solutions exist globally in time and remain uniformly bounded; Lyapunov functionals prove that the spatially homogeneous prey-only and coexistence equilibria are globally asymptotically stable under suitable parameter restrictions.
What carries the argument
Lyapunov functionals constructed from the system with prey-density-dependent motility and mobility coefficients
If this is right
- Global existence and uniform boundedness hold for the classical solutions in two dimensions.
- The prey-only equilibrium is globally asymptotically stable for small predator growth rates or large death rates.
- The coexistence equilibrium is globally asymptotically stable when the motility function satisfies a monotonicity condition and parameters lie in a stated range.
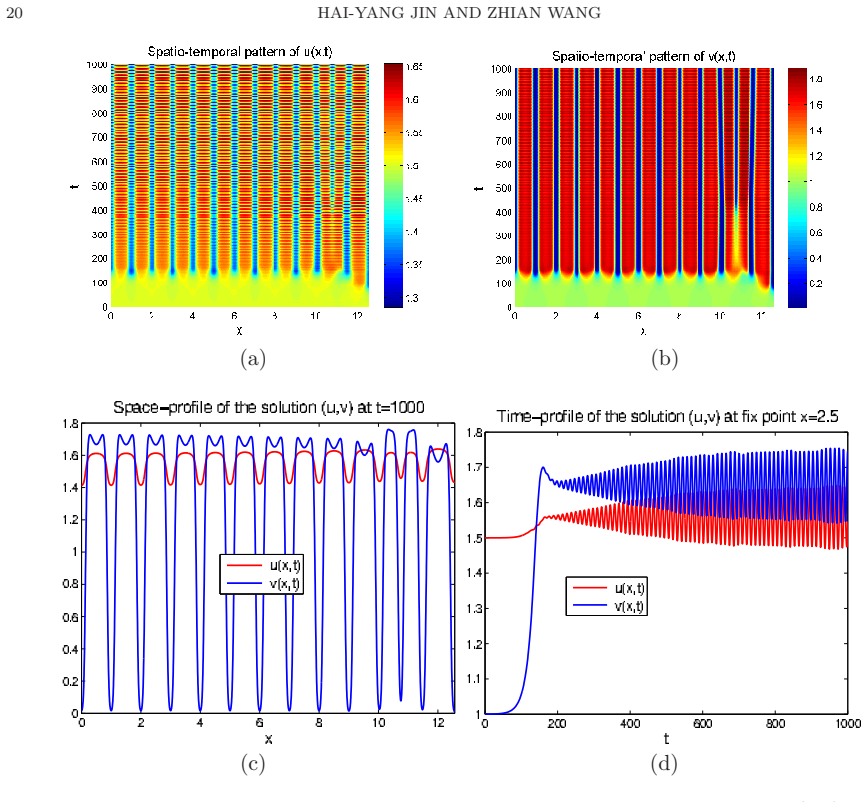
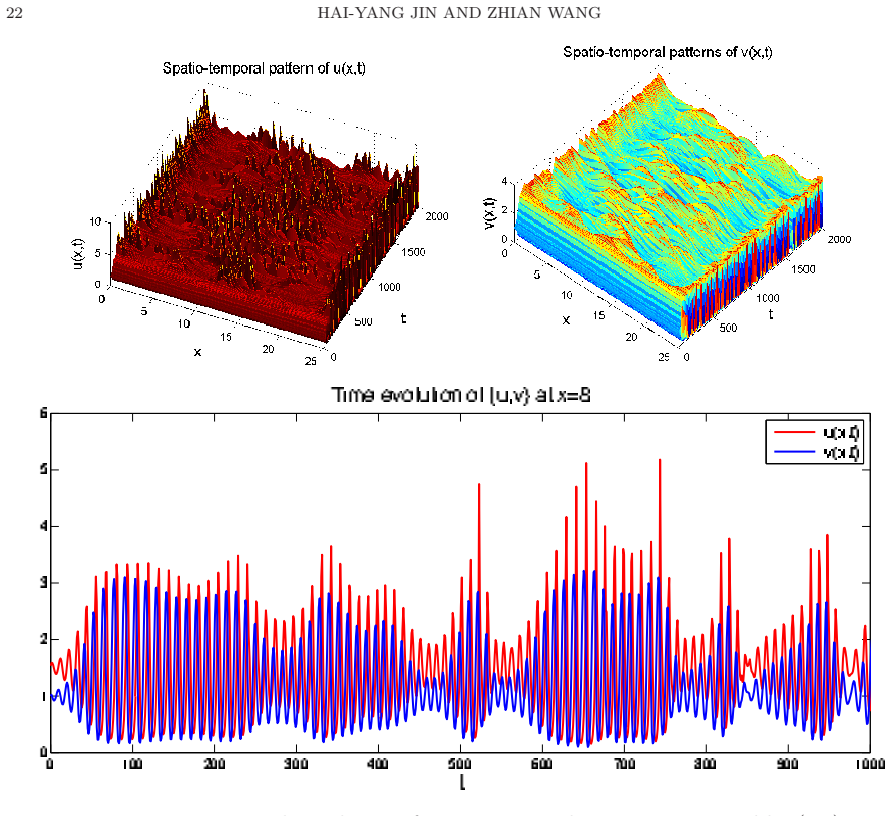
- Outside the stability regime, the model produces time-periodic, stationary inhomogeneous, or chaotic spatio-temporal patterns.
Where Pith is reading between the lines
- The explicit linkage between motility and taxis coefficients may be the mechanism that prevents finite-time blow-up common in other taxis models.
- The observed mismatch between linearized and fully nonlinear temporal dynamics implies that pattern selection is governed by nonlinear terms rather than linear instability alone.
- The same motility function could be tested in three-dimensional domains or with different boundary conditions to check whether boundedness persists.
Load-bearing premise
The predator's motility and mobility coefficients are linked by a prey-density-dependent function that satisfies the technical conditions required for the Lyapunov construction and the boundedness argument.
What would settle it
Explicit construction of a solution that becomes unbounded in finite time, or direct numerical simulation showing that one of the homogeneous equilibria loses stability while the Lyapunov conditions still hold.
Figures

read the original abstract
In this paper, we investigate the global boundedness, asymptotic stability and pattern formation of predator-prey systems with density-dependent preytaxis in a two-dimensional bounded domain with Neumann boundary conditions, where the coefficients of motility (diffusion) and mobility (preytaxis) of the predator are correlated through a prey density dependent motility function. We establish the existence of classical solutions with uniform-in time bound and the global stability of the spatially homogeneous prey-only steady states and coexistence steady states under certain conditions on parameters by constructing Lyapunov functionals. With numerical simulations, we further demonstrate that spatially homogeneous time-periodic patterns, stationary spatially inhomogeneous patterns and chaotic spatio-temporal patterns are all possible for the parameters outside the stability regime. We also find from numerical simulations that the temporal dynamics between linearized system and nonlinear systems are quite different, and the prey density-dependent motility function can trigger the pattern formation.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper investigates a quasilinear parabolic predator-prey system in a bounded 2D domain with Neumann BCs, where predator motility and preytaxis coefficients are linked via a prey-density-dependent function m(v). It claims to establish global existence of classical solutions that remain uniformly bounded in time, together with asymptotic stability of the spatially homogeneous prey-only and coexistence equilibria, under suitable parameter restrictions, via construction of Lyapunov functionals. Numerical simulations are presented to show that time-periodic, stationary inhomogeneous, and chaotic patterns can occur outside the stability regime, and that the motility function can induce pattern formation.
Significance. If the proofs are complete, the results would contribute to the analysis of taxis-diffusion systems by showing how density-dependent motility can yield global boundedness and stability where standard taxis models may not, while the numerics illustrate the transition to complex dynamics. The explicit construction of Lyapunov functionals from the model equations is a strength when the technical conditions on m(v) hold.
major comments (2)
- [Section 2] Section 2 (model assumptions): the boundedness and stability proofs rely on technical conditions on the motility function m(v) (positivity, monotonicity, and growth restrictions) that allow absorption of cross terms in the energy estimates and closure of the L^∞ bounds; these conditions are not shown to hold for arbitrary biologically plausible choices of m(v), and the paper does not provide concrete examples or discuss the consequences when they fail.
- [Introduction / abstract] The abstract and introduction claim global stability of both prey-only and coexistence steady states 'under certain conditions on parameters' via Lyapunov functionals, but the precise parameter restrictions and the explicit form of the functionals (including how the m(v)-dependent terms are controlled) are not stated in a way that allows verification of the dissipation estimate without the full proofs.
minor comments (1)
- [Abstract] Abstract: 'uniform-in time' should read 'uniform-in-time'.
Simulated Author's Rebuttal
We thank the referee for the careful reading of our manuscript and the constructive comments. We provide point-by-point responses to the major comments below. We will revise the manuscript to address the issues raised.
read point-by-point responses
-
Referee: [Section 2] Section 2 (model assumptions): the boundedness and stability proofs rely on technical conditions on the motility function m(v) (positivity, monotonicity, and growth restrictions) that allow absorption of cross terms in the energy estimates and closure of the L^∞ bounds; these conditions are not shown to hold for arbitrary biologically plausible choices of m(v), and the paper does not provide concrete examples or discuss the consequences when they fail.
Authors: The conditions on the motility function m(v) are necessary to control the cross-diffusion terms in the energy estimates and to obtain the uniform L^∞ bounds. We agree that concrete examples would be helpful. In the revised manuscript, we will add specific examples of functions m(v) that satisfy all the required assumptions (e.g., m(v) = 1/(1 + v) which is positive, decreasing, and satisfies the growth bound). We will also add a short discussion noting that if the growth restrictions on m(v) are violated, the current proof technique does not apply and global boundedness may fail, although this is outside the scope of the present work. revision: yes
-
Referee: [Introduction / abstract] The abstract and introduction claim global stability of both prey-only and coexistence steady states 'under certain conditions on parameters' via Lyapunov functionals, but the precise parameter restrictions and the explicit form of the functionals (including how the m(v)-dependent terms are controlled) are not stated in a way that allows verification of the dissipation estimate without the full proofs.
Authors: While the detailed conditions and functionals are presented in the theorems and proofs in Sections 3 and 4, we acknowledge that the abstract and introduction could better highlight them for the reader. In the revision, we will expand the introduction to explicitly list the main parameter conditions (such as those ensuring the dissipation of the Lyapunov functional) and briefly describe how the m(v)-dependent terms are handled in the estimates. revision: yes
Circularity Check
No significant circularity; proofs are self-contained under explicit assumptions
full rationale
The central results rely on direct construction of Lyapunov functionals from the PDE system itself (as described in the abstract and section 2), with explicit technical assumptions on the motility function m(v) stated upfront to close the estimates. These assumptions are not derived from the stability conclusions but are independent conditions under which the dissipation works. Numerical patterns are reported as observations from simulations outside the analytically stable regime, with no fitted parameters renamed as predictions and no load-bearing self-citations reducing the derivation to prior author work. The chain does not reduce to its inputs by construction.
Axiom & Free-Parameter Ledger
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
We establish the existence of classical solutions with uniform-in-time bound and the global stability … by constructing Lyapunov functionals.
-
IndisputableMonolith/Foundation/AlphaCoordinateFixation.leanalpha_pin_under_high_calibration unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
If χ(v)=−d′(v), the system becomes ut=Δ(d(v)u)+… (density-suppressed motility)
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
Alikakos, Lp bounds of solutions of reaction-diffusion equations
N.D. Alikakos, Lp bounds of solutions of reaction-diffusion equations. Comm. Partial Dif- ferential Equations, 4:827-868, 1979
work page 1979
-
[2]
B.E. Ainseba, M. Bendahmane, and A. Noussair, A reaction -diffusion system modeling predator-prey with prey-taxis. Nonlinear Anal. Real World Appl., 9(5):2086-2105, 2008. GLOBAL STABILITY AND SPATIO-TEMPORAL PATTERNS OF PREDATOR -PREY SYSTEMS 23
work page 2086
-
[3]
Amann, Dynamic theory of quasilinear parabolic equat ions III
H. Amann, Dynamic theory of quasilinear parabolic equat ions III. Global existence. Math. Z., 202:219-250, 1989
work page 1989
-
[4]
Amann, Dynamic theory of quasilinear parabolic equat ions II
H. Amann, Dynamic theory of quasilinear parabolic equat ions II. Reaction-diffusion sys- tems. Differ. Integral Equ., 3(1):13-75, 1990
work page 1990
-
[5]
Amann, Nonhomogeneous linear and quasilinear ellipt ic and parabolic boundary value problems
H. Amann, Nonhomogeneous linear and quasilinear ellipt ic and parabolic boundary value problems. In Function spaces, differential operators and non linear analysis (Friedrichroda, 1992), volume 133 of Teubner-Texte Math., pages 9-126. Teub ner, Stuttgart, 1993
work page 1992
-
[6]
A. Chakraborty, M. Singh, D. Lucy, and P. Ridland, Predat or-prey model with prey-taxis and diffusion. Math. Comp. Mod ., 46:482-498, 2007
work page 2007
-
[7]
P. Chesson and W. Murdoch, Aggregation of risk: relation ships among host-parasitoid medels. Am. Nat., 127:696-715, 1986
work page 1986
-
[8]
Cosner, Reaction-diffusion-advection models for the e ffects and evolution of dispersal
C. Cosner, Reaction-diffusion-advection models for the e ffects and evolution of dispersal. Discrere. Contin. Dyn. Syst., 34:1701-1745, 2014
work page 2014
-
[9]
E. Curio. The Ethology of Predation . Spring -Verlag, New York, 1976
work page 1976
- [10]
-
[11]
D. Gr¨ unbaum, Using spatially explicit models to chara cterize foraging performance in het- erogeneous landscapes. Am. Nat ., 151:97-115, 1998
work page 1998
-
[12]
C.S. Holling, The functional response of predators to p rey density and its role in mimicry and population regulation. Mem. Entom. Soc. Can., 45:1-60, 1965
work page 1965
-
[13]
H.Y. Jin and Z.A. Wang, Global stability of prey-taxis s ystems. J. Differential Equations, 262:1257-1290, 2017
work page 2017
- [14]
-
[15]
J¨ ungel,Diffusive and Nondiffusive Population Models
A. J¨ ungel,Diffusive and Nondiffusive Population Models. Mathematical modeling of collec- tive behavior in socio-economic and life sciences , 397-425, Model. Simul. Sci. Eng. Technol., Birkh¨ auser Boston, Inc., Boston, MA, 2010
work page 2010
-
[16]
A. J¨ ungel, C. Kuehn and L. Trussardi, A meeting point of entr opy and bifurcations in cross-diffusion herding. European J. Appl. Math., 28(2):317-356, 2017
work page 2017
- [17]
-
[18]
E.F. Keller and L.A. Segel, Model for chemotaxis. J. Theor. Biol. , 30(2):225-234,1971
work page 1971
-
[19]
W.G. Kelley and A.C. Peterson, The Thoery of Differential Equations - Classical and Qual- itative, Springer, 2010
work page 2010
-
[20]
R. Kowalczyk and Z. Szyma´ nska, On the global existence of solutions to an aggregation model. J. Math. Anal. Appl., 343:379-398, 2008
work page 2008
-
[21]
O. Ladyzhenskaya, V. Solonnikov, and N. Uralceva, Linear and Quasilinear Equations of Parabolic Type. AMS, Providence, RI, 1968
work page 1968
-
[22]
LaSalle, Some extensions of Lyapunov’s second met hod
J.P. LaSalle, Some extensions of Lyapunov’s second met hod. IRE Transactions on Circuit Theory, CT-7 , 520-527, 1960
work page 1960
-
[23]
J.M. Lee, T. Hillen, and M.A. Lewis, Continuous travlin g waves for prey-taxis. Bull. Math. Biol., 70:654-676, 2008
work page 2008
-
[24]
J.M. Lee, T. Hillen, and M.A. Lewis, Pattern formation i n prey-taxis systems. J. Biol. Dyn., 3(6):551-573, 2009
work page 2009
- [25]
-
[26]
Lotka, Elements of Physical Biology
A.J. Lotka, Elements of Physical Biology . Baltimore: Williams and Wilkins Co., 1925
work page 1925
-
[27]
M. Mimura and K. Kawasaki, Spatial segregation in compe titive interaction-diffusion equa- tions. J. Math. Biol., 9(1):49-64, 1980. 24 HAI-YANG JIN AND ZHIAN W ANG
work page 1980
-
[28]
N. Mizoguchi and P. Souplet, Nondegeneracy of blow-up p oints for the parabolic Keller- Segel system. Ann. Inst. H. Poincar´ e Anal. Non Lin´ eaire,31:851-875, 2014
work page 2014
-
[29]
W.W. Murdoch, C.J. Briggs, and R.M. Nisbert, Consumer-Resource Dynamics (Mono- graphs in Population Biology-36) . Princeton University Press, 2003
work page 2003
-
[30]
W. Murdoch, J. Chesson, and P. Chesson, Biological cont rol in theory and practice. Am. Nat., 125:344-366, 1985
work page 1985
-
[31]
A. Okubo and S.A. Levin, Diffusion and Ecological Problems: Modern Perspective . Inter- disciplinary Applied Mathematics, vol. 14. 2nd ed. Berlin: Springer, 2001
work page 2001
-
[32]
M.M. Porzio and V.Vespri, H¨ older estimates for local s olutions of some doubly nonlinear degenerate parabolic equations. J. Differential Equations, 103(1):146-178, 1993
work page 1993
-
[33]
M.L. Rosenzweig and R.H. MacArthur, Graphical represe ntation and stability conditions of predator-prey interactions. Am. Nat ., 97:209-223, 1963
work page 1963
-
[34]
N. Sapoukhina, Y. Tyutyunov, and R. Arditi, The role of p rey taxis in biological control: a spatial theoretical model. Am. Nat ., 162:61-76, 2003
work page 2003
-
[35]
J. Smith-Roberge, D. Iron and T. Kolokolnikov, Pattern formation in bacterial colonies with density-dependent diffusion. European J. Appl. Math. , 30(1):196-218, 2019
work page 2019
-
[36]
P. Souplet and P. Quittner, Superlinear Parabolic Problems: Blow-up, Global Existenc e and Steady States. Birkh¨ auser Advanced Texts, Basel/Boston/Berlin, 2007
work page 2007
-
[37]
Tao, Global existence of classical solutions to a p redator-prey model with nonlinear prey-taxis
Y.S. Tao, Global existence of classical solutions to a p redator-prey model with nonlinear prey-taxis. Nonlinear Anal. Real World Appl., 11(3):2056-2064, 2010
work page 2056
- [38]
- [39]
-
[40]
P. Turchin, Complex Population Dynamics: A Theoretical/Empirical Synth esis (Mono- graphs in Population Biology-35) . Princeton University Press, 2003
work page 2003
-
[41]
Volterra, Fluctuations in the abundance of a species considered mathematically
V. Volterra, Fluctuations in the abundance of a species considered mathematically. Nature, 118:558-560, 1926
work page 1926
-
[42]
K. Wang, Q. Wang and F. Yu, Stationary and time periodic p atterns of two predator and one-prey systems with prey-taxis. Discrere. Contin. Dyn. Syst., 37(1):505-543, 2017
work page 2017
-
[43]
Wang, Note on the Lyapunov functional method
M.X. Wang, Note on the Lyapunov functional method. Appl. Math. Lett., 75:102-107, 2018
work page 2018
-
[44]
Q. Wang, Y. Song and L.J. Shao, Nonconstant positive ste ady states and pattern formation of 1D prey-taxis systems. J. Nonlinear Sci., 27(1):71-97, 2017
work page 2017
-
[45]
J.P. Wang and M.X.Wang, Boundedness and global stabili ty of the two-predator and one- prey models with nonlinear prey-taxis. Z. Angew. Math. Phys., 69(3): 63, 24pp, 2018
work page 2018
-
[46]
S. Wu, J.P. Shi, and B. Wu, Global existence of solutions and uniform persistence of a diffusive predator-prey model with prey-taxis. J. Differential Equations , 260(7):5847-5874, 2016
work page 2016
-
[47]
S. Wu, J Wang and J. Shi, Dynamics and pattern formation o f a diffusive predator-prey model with predator-taxis. Math. Models Method Appl. Sci., 28(11):2275-2312, 2018
work page 2018
-
[48]
F. Yi, J. Wei and J. Shi, Bifurcation and spatiotemporal patterns in a homogeneous diffusive predator-prey system, J. Differential Equations , 246:1944-1977, 2009. Department of Mathematics, South China University of Techn ology, Guangzhou 510640, China E-mail address : mahyjin@scut.edu.cn Department of Applied Mathematics, Hong Kong Polytechnic U niversity...
work page 1944
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.