Cell bulging and extrusion in a three-dimensional bubbly vertex model for curved epithelial sheets
Pith reviewed 2026-05-23 17:29 UTC · model grok-4.3
The pith
Epithelial curvature leads to cell bulging and extrusion at topological defects through amplified interfacial forces.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
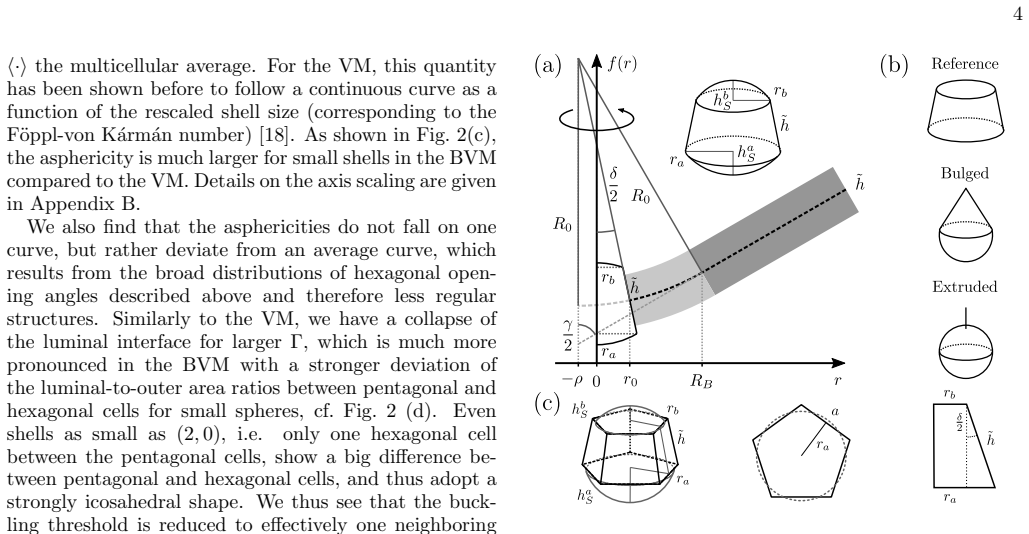
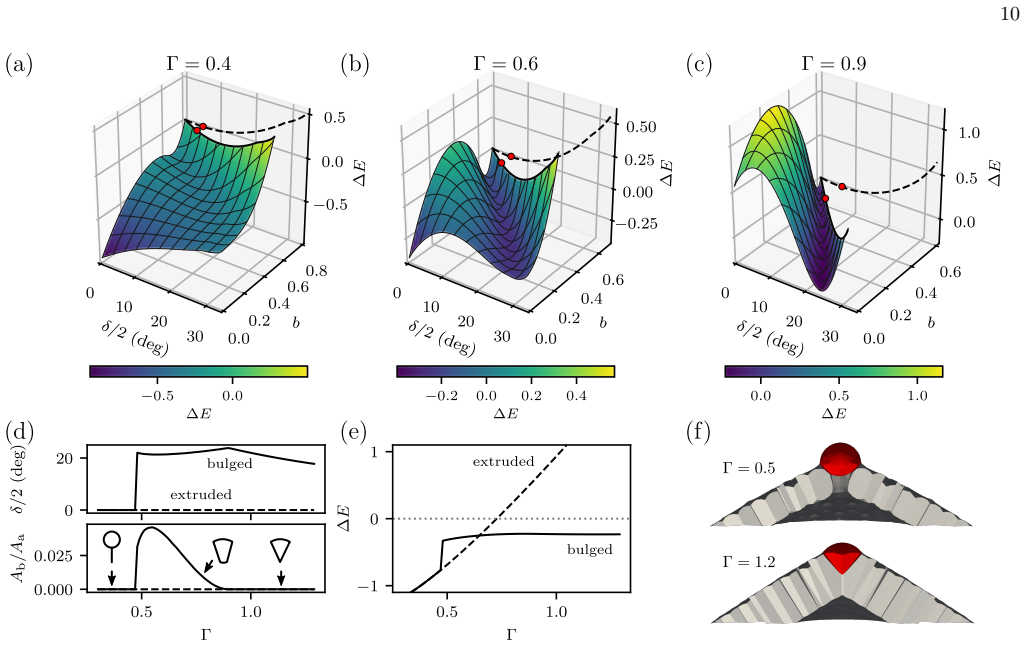
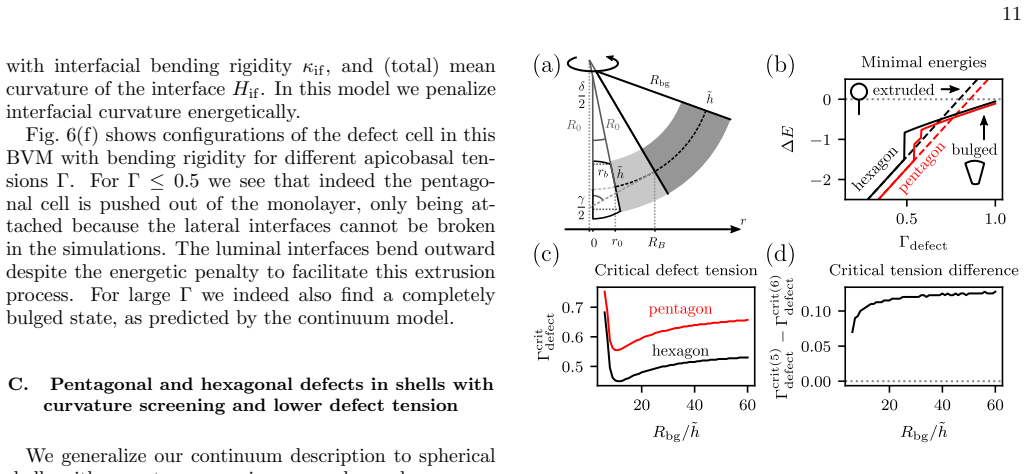
In the three-dimensional bubbly vertex model, topological defects in curved epithelial sheets trigger a cellular bulging instability because the interfacial curvature at individual cells strongly amplifies the buckling induced by tissue-scale defects. Increased basal interfacial tension in pentagonal cells, inferred from organoid imaging, stabilizes against this instability and achieves superior cell shape control.
What carries the argument
three-dimensional bubbly vertex model with basal interfacial tension that amplifies defect-induced buckling via cell interface curvature
If this is right
- Curved epithelial tissues naturally develop bulged and extrusion-like cell shapes at topological defects.
- Basal tension adjustments in pentagonal cells provide effective local stabilization of cell shapes.
- Tissue-scale mechanisms such as lumen pressure or spontaneous curvature are less effective for shape control than local basal tension.
- Extrusion serves as a density control mechanism that is enhanced by curvature in three-dimensional sheets.
Where Pith is reading between the lines
- The same amplification mechanism may operate in other curved epithelia such as lung or intestinal tissues.
- Direct tension measurements at defects across varying curvatures could test the inferred cellular response.
- Topological defects likely influence 3D tissue organization more strongly than in flat monolayers.
Load-bearing premise
Increased basal interfacial tension in pentagonal cells inferred from 3D imaging of spherical mouse colon organoids accurately reflects and counters the force conditions at topological defects.
What would settle it
Direct observation of no increased basal tension specifically in pentagonal cells at defects in curved epithelia, or persistent bulging instability despite such tension, would falsify the proposed stabilization.
Figures






read the original abstract
Cell extrusion is an essential mechanism for controlling cell density in epithelial tissues. Another essential element of epithelia is curvature, which is required to achieve complex shapes, like in the lung or intestine. Here we introduce a three-dimensional bubbly vertex model to study the interplay between extrusion and curvature. We find a generic cellular bulging instability at topological defects which is much stronger than for standard vertex models. Analyzing cell shapes in three-dimensional imaging data of spherical mouse colon organoids, we infer that pentagonal cells have an increased basal interfacial tension, suggesting that cells at topological defects react to the different force conditions. Using the bubbly vertex model, we show that such basal tensions stabilize against the predicted instability and result in better cell shape control than tissue-scale mechanisms such as lumen pressure and spontaneous curvature. Our theory suggests that epithelial curvature naturally leads to bulged and extrusion-like cell shapes because the interfacial curvature of individual cells at the defects strongly amplifies buckling effected by tissue-scale topological defects in elastic sheets. Our results highlight the complex interplay of forces across scales in three-dimensional tissue organization.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript introduces a three-dimensional bubbly vertex model for curved epithelial sheets and reports a generic cellular bulging instability at topological defects that is amplified by individual-cell interfacial curvature, stronger than in standard vertex models. From 3D imaging of spherical mouse colon organoids the authors infer increased basal interfacial tension in pentagonal cells. The model is then used to show that this tension stabilizes the instability more effectively than tissue-scale mechanisms such as lumen pressure or spontaneous curvature. The central theoretical claim is that epithelial curvature naturally produces bulged and extrusion-like shapes because cell interfacial curvature at defects amplifies buckling driven by tissue-scale topological defects.
Significance. If the central claims are substantiated, the work would be significant for understanding multi-scale force balance in 3D epithelial morphogenesis, particularly the interplay between topology, curvature, and cell extrusion. The development of the bubbly vertex model and its direct comparison to organoid imaging data constitute clear strengths. The result offers a mechanistic explanation that could be tested in other curved epithelia.
major comments (1)
- [Section describing organoid imaging analysis and tension inference] The mapping from organoid shape statistics to an increased basal interfacial tension specifically at pentagonal cells (and its identification as the biologically relevant counter-force at topological defects) is not secured by controls for global curvature, lumen pressure, or cell-density effects. This assumption is load-bearing for the claim that the inferred tension stabilizes the instability better than tissue-scale mechanisms, yet the experimental section provides no direct validation that the observed pentagon bias encodes the local force imbalance present in the model.
minor comments (1)
- Figure captions should explicitly state the number of organoids and cells analyzed and include scale bars; model parameter tables would benefit from a column listing which parameters are fixed versus fitted.
Simulated Author's Rebuttal
We thank the referee for their constructive review and for highlighting the need to strengthen the experimental inference section. We respond to the single major comment below.
read point-by-point responses
-
Referee: [Section describing organoid imaging analysis and tension inference] The mapping from organoid shape statistics to an increased basal interfacial tension specifically at pentagonal cells (and its identification as the biologically relevant counter-force at topological defects) is not secured by controls for global curvature, lumen pressure, or cell-density effects. This assumption is load-bearing for the claim that the inferred tension stabilizes the instability better than tissue-scale mechanisms, yet the experimental section provides no direct validation that the observed pentagon bias encodes the local force imbalance present in the model.
Authors: We acknowledge that the inference of elevated basal tension in pentagons rests on statistical differences in cell shape between pentagons and hexagons without dedicated controls that isolate global curvature, lumen pressure, or density. Global parameters would be expected to affect all cells similarly and therefore cannot account for the topology-specific bias we report. We will add a new paragraph in the revised manuscript that explicitly discusses these assumptions, notes the absence of direct force measurements, and clarifies that the model comparison demonstrates consistency with local tension adjustment rather than tissue-scale alternatives. This constitutes a partial revision focused on improved transparency rather than new data. revision: partial
Circularity Check
No circularity: model predictions independent of fitted inputs; imaging inference treated as external constraint
full rationale
The derivation introduces a new 3D bubbly vertex model, derives a bulging instability at defects from its equations, then uses independent 3D organoid imaging to infer a basal-tension adjustment that is inserted as a parameter. No step reduces a claimed prediction to a fit by construction, no self-citation chain bears the central result, and the imaging-to-tension mapping is presented as an external observation rather than derived from the model itself. The overall claim therefore rests on the model's independent mechanics plus separate data, not on re-labeling of its own inputs.
Axiom & Free-Parameter Ledger
free parameters (1)
- basal interfacial tension for pentagonal cells
axioms (1)
- domain assumption The 3D bubbly vertex model appropriately captures mechanics of curved epithelial sheets including curvature and extrusion
Lean theorems connected to this paper
-
IndisputableMonolith/Foundation/AlexanderDuality.leanalexander_duality_circle_linking echoes?
echoesECHOES: this paper passage has the same mathematical shape or conceptual pattern as the Recognition theorem, but is not a direct formal dependency.
We find a generic cellular bulging instability at topological defects which is much stronger than for standard vertex models... extrusion is energetically preferred at topological defects due to a redistribution of Gaussian curvature into the defect.
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
the total energy of the tissue is then described by the sum of the individual surface area contributions E = Σ_i (Γ_a A_a,i + Γ_b A_b,i + ½ Γ_l A_l,i)
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
Bending energies For the cap segment we consider the radial deflection function f with rotational symmetry f(r) = R0 − q R2 0 − (ρ + r)2, (8) where ρ is the radial offset due to the bulged defect cell, cf. Fig. 3. The bending energy density reads ebending = κ 2 H2 + κGK, (9) with bending rigidity κ, saddle splay modulus κG, (total) mean curvature H = c+ c...
-
[2]
Stretching energies The stretching energy density reads estretch = 1 2 2µε2 ij + λε2 kk , (15) with two-dimensional Lam´ e coefficientsµ and λ. To com- pute the stretching energy of the tissue we consider the in-plane deformation of a conically bent elastic sheet. Using polar coordinates ( r, θ), it is clear from symme- try that the azimuthal deformation ...
-
[3]
Since we only consider a linear f, i.e
This results in the corrected radial strain εrr = w′(r) −(1+ f1 2)−1/2. Since we only consider a linear f, i.e. we neglect f ′′, only the term corresponding to ∆s changes in Eq. (17), implying that the solution is again Eq. (18), now with ∆s = qs − 1 − 1p 1 + f1 2 ! . (19) As boundary conditions we assume vanishing radial stresses at the inner and outer r...
-
[4]
G. T. Eisenhoffer, P. D. Loftus, M. Yoshigi, H. Otsuna, C.-B. Chien, P. A. Morcos, and J. Rosenblatt, Nature 484, 546 (2012)
work page 2012
-
[5]
S. A. Gudipaty, J. Lindblom, P. D. Loftus, M. J. Redd, K. Edes, C. F. Davey, V. Krishnegowda, and J. Rosen- blatt, Nature 543, 118 (2017)
work page 2017
-
[6]
S. A. Gudipaty and J. Rosenblatt, Semin. Cell Dev. Biol. 67, 132 (2017)
work page 2017
-
[7]
L. Kocgozlu, T. Saw, A. Le, I. Yow, M. Shagirov, E. Wong, R.-M. M` ege, C. Lim, Y. Toyama, and B. Ladoux, Curr. Biol. 26, 2942 (2016)
work page 2016
-
[8]
A. P. Le, J.-F. Rupprecht, R.-M. M` ege, Y. Toyama, C. T. Lim, and B. Ladoux, Nat. Commun. 12, 397 (2021)
work page 2021
-
[9]
L. G. van der Flier and H. Clevers, Annu. Rev. Physiol. 71, 241 (2009)
work page 2009
-
[10]
C. P´ erez-Gonz´ alez, G. Ceada, M. Matejˇ ci´ c, and X. Trepat, Curr. Opin. Genet. Dev. 72, 82 (2022)
work page 2022
-
[11]
T. B. Saw, A. Doostmohammadi, V. Nier, L. Kocgozlu, S. Thampi, Y. Toyama, P. Marcq, C. T. Lim, J. M. Yeo- mans, and B. Ladoux, Nature 544, 212 (2017)
work page 2017
- [12]
-
[13]
T. Chen, T. B. Saw, R.-M. M` ege, and B. Ladoux, J. Cell Sci. 131, jcs218156 (2018)
work page 2018
-
[14]
W. Tang, A. Das, A. F. Pegoraro, Y. L. Han, J. Huang, D. A. Roberts, H. Yang, J. J. Fredberg, D. N. Kotton, D. Bi, and M. Guo, Nat. Phys. 18, 1371 (2022)
work page 2022
-
[15]
L. A. Hoffmann, L. N. Carenza, J. Eckert, and L. Giomi, Sci. Adv. 8, eabk2712 (2022)
work page 2022
- [16]
-
[17]
J.-M. Armengol-Collado, L. N. Carenza, J. Eckert, D. Krommydas, and L. Giomi, Nat. Phys. 19, 1773 (2023)
work page 2023
-
[18]
S. Monfared, G. Ravichandran, J. Andrade, and A. Doostmohammadi, eLife 12, e82435 (2023)
work page 2023
-
[19]
C. Bielmeier, S. Alt, V. Weichselberger, M. La Fortezza, H. Harz, F. J¨ ulicher, G. Salbreux, and A.-K. Classen, Curr. Biol. 26, 563 (2016)
work page 2016
- [20]
-
[21]
O. M. Drozdowski and U. S. Schwarz, Phys. Rev. Res. 6, L022045 (2024)
work page 2024
- [22]
-
[23]
T. T. Nguyen, R. F. Bruinsma, and W. M. Gelbart, Phys. Rev. E 72, 051923 (2005)
work page 2005
-
[24]
T. A. Witten and H. Li, EPL 23, 51 (1993)
work page 1993
-
[25]
H. S. Seung and D. R. Nelson, Phys. Rev. A 38, 1005 (1988)
work page 1988
- [26]
-
[27]
A. Boromand, A. Signoriello, F. Ye, C. S. O’Hern, and M. D. Shattuck, Phys. Rev. Lett. 121, 248003 (2018)
work page 2018
- [28]
- [29]
- [30]
- [31]
-
[32]
O. M. Drozdowski and U. S. Schwarz, “OrganoidChaste: A three-dimensional vertex model for epithelial mono- layers (WILL BE PUBLISHED UPON FINAL ACCEP- TANCE),” https://github.com/oliverdrozdowski/ OrganoidChaste (2024)
work page 2024
-
[33]
F. R. Cooper, R. E. Baker, M. O. Bernabeu, R. Bordas, L. Bowler, A. Bueno-Orovio, H. M. Byrne, V. Carapella, L. Cardone-Noott, J. Cooper, S. Dutta, B. D. Evans, A. G. Fletcher, J. A. Grogan, W. Guo, D. G. Harvey, M. Hendrix, D. Kay, J. Kursawe, P. K. Maini, B. McMil- lan, G. R. Mirams, J. M. Osborne, P. Pathmanathan, J. M. Pitt-Francis, M. Robinson, B. Ro...
work page 2020
-
[34]
D. S. Roshal, K. Azzag, K. K. Fedorenko, S. B. Rochal, and S. Baghdiguian, Phys. Rev. E 108, 024404 (2023)
work page 2023
-
[35]
K. A. Brakke, Exp. Math. 1, 141 (1992)
work page 1992
-
[36]
I. Garc´ ıa-Aguilar, P. Fonda, and L. Giomi, Phys. Rev. E 101, 063005 (2020)
work page 2020
-
[37]
D. L. D. Caspar and A. Klug, Cold Spring Harb. Symp. Quant. Biol. 27, 1 (1962)
work page 1962
-
[38]
L. D. Landau and E. M. Lifshitz, Theory of elasticity , 2nd ed., Course of theoretical physics, Vol. 7 (Pergamon Press, Oxford, 1970)
work page 1970
-
[39]
Q. Yang, S.-L. Xue, C. J. Chan, M. Rempfler, D. Vis- chi, F. Maurer-Gutierrez, T. Hiiragi, E. Hannezo, and P. Liberali, Nat. Cell Biol. 23, 733 (2021)
work page 2021
-
[40]
E. Hannezo, J. Prost, and J.-F. Joanny, Proc. Natl. Acad. Sci. U.S.A. 111, 27 (2014)
work page 2014
- [41]
-
[42]
N. Gjorevski, M. Nikolaev, T. E. Brown, O. Mitrofanova, N. Brandenberg, F. W. DelRio, F. M. Yavitt, P. Liberali, K. S. Anseth, and M. P. Lutolf, Science 375, eaaw9021 (2022)
work page 2022
-
[43]
J. Y. Co, M. Margalef-Catal` a, X. Li, A. T. Mah, C. J. Kuo, D. M. Monack, and M. R. Amieva, Cell Rep. 26, 2509 (2019)
work page 2019
-
[44]
J. Y. Co, M. Margalef-Catal` a, D. M. Monack, and M. R. Amieva, Nat. Protoc. 16, 5171 (2021)
work page 2021
- [45]
-
[46]
M. M. Terzi and M. Deserno, J. Chem. Phys.147, 084702 (2017)
work page 2017
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.