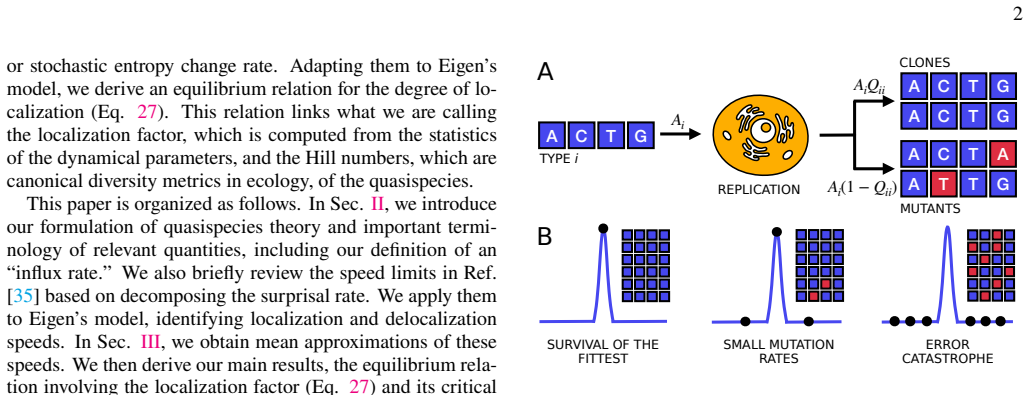
Quasilocalization under coupled mutation-selection dynamics
Pith reviewed 2026-05-15 21:41 UTC · model grok-4.3
The pith
A general relation links quasispecies Hill numbers to the ratio of effective fitness variance over mean mutation rate squared.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
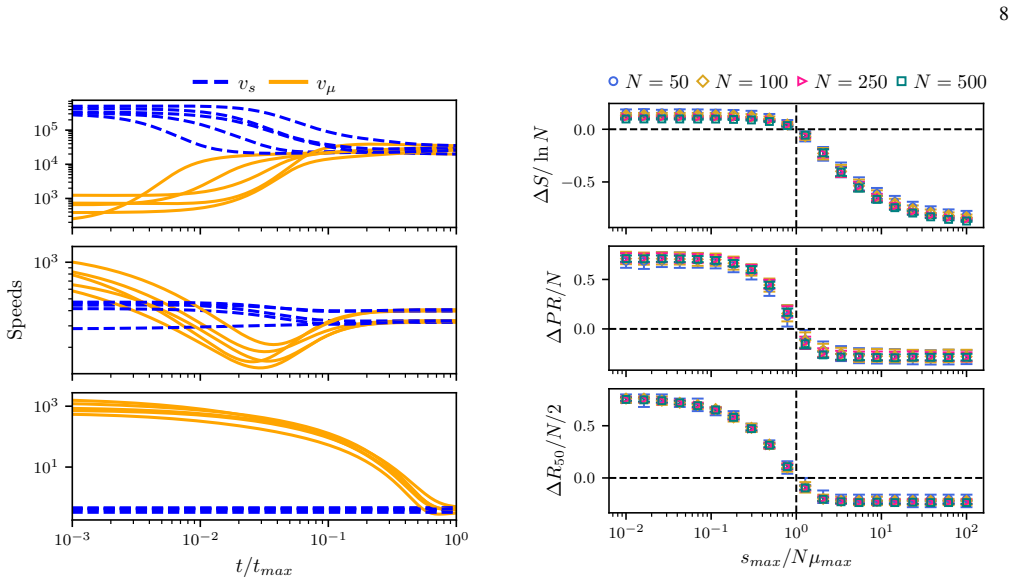
Under coupled mutation-selection dynamics, the Hill numbers of the quasispecies distribution satisfy a direct relation to the localization factor, given by the ratio of an effective fitness variance to the mean mutation rate squared. The relation follows from mean approximations applied to the decomposed rates of surprisal or stochastic entropy change.
What carries the argument
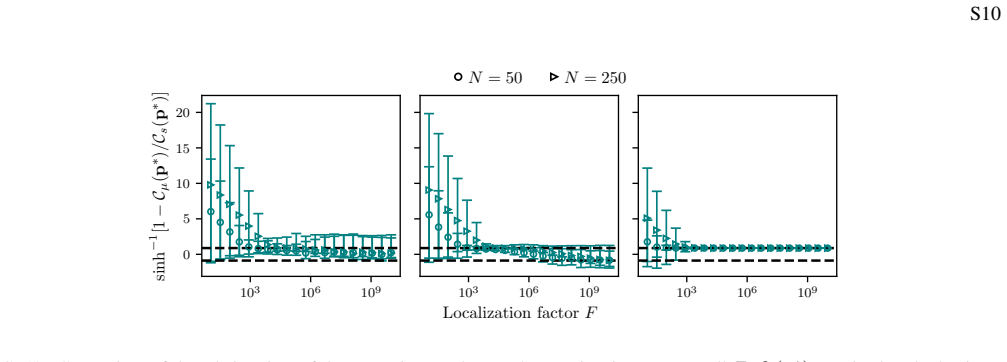
The localization factor, the ratio of effective fitness variance to the square of the mean mutation rate, which sets the values taken by the quasispecies Hill numbers via averaged surprisal or entropy change rates.
If this is right
- Localization of quasispecies populations can be estimated for any fitness landscape using the localization factor without full stochastic simulation.
- Combinations of Hill numbers acquire a dynamical meaning and can serve as complexity measures for real viral sequence data under Eigen's model.
- The relation supplies a quantitative description of delocalization that occurs beyond critical mutation thresholds.
- Effective fitness variance can be inferred from observed Hill numbers once mutation rates are known.
Where Pith is reading between the lines
- Hill numbers extracted from viral sequencing data could be inverted to estimate hidden effective fitness variances in natural populations.
- The approximation might be tested for robustness in finite-population or spatially structured versions of the model.
- The link between ecological diversity indices and evolutionary entropy rates could be checked for consistency with other population-genetic entropy measures.
- Similar relations might appear in non-viral systems such as bacterial populations under changing selection pressures.
Load-bearing premise
The mean approximations of the decomposed surprisal or stochastic entropy change rates remain valid for arbitrary fitness landscapes and mutation rates.
What would settle it
In an experimental viral population with a known fitness landscape, measure the Hill numbers at controlled mutation rates, compute the implied localization factor, and check whether it equals the independently measured ratio of effective fitness variance to mutation rate squared.
Figures






read the original abstract
When mutations are rampant, quasispecies theory or Eigen's model predicts that the fittest type in a population may not dominate. Beyond a critical mutation rate, the population may even be delocalized completely from the peak of the fitness landscape and the fittest is ironically lost. Extensive efforts have been made to understand this exceptional scenario. But in general, there is no simple prescription that predicts the eventual degree of localization for arbitrary fitness landscapes and mutation rates. Here, we derive a simple and general relation linking the quasispecies' Hill numbers, which are diversity metrics in ecology, and the ratio of an effective fitness variance to the mean mutation rate squared. This ratio, which we call the localization factor, emerges from mean approximations of decomposed surprisal or stochastic entropy change rates. On the side of application, the relation we obtained here defines a combination of Hill numbers that may complement other complexity or diversity measures for real viral quasispecies. Its advantage being that there is an underlying biological interpretation under Eigen's model.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript derives a relation linking quasispecies Hill numbers (as diversity metrics) to a localization factor given by the ratio of effective fitness variance to the square of the mean mutation rate. The relation is obtained from mean approximations applied to decomposed surprisal or stochastic entropy change rates within Eigen's coupled mutation-selection model, and is proposed as a general, biologically interpretable descriptor of localization for arbitrary fitness landscapes.
Significance. If the mean approximations hold with controllable error, the result supplies a parameter-light, model-grounded combination of Hill numbers that directly quantifies localization degree. This could complement existing complexity measures for viral quasispecies data and offers a falsifiable link between observable diversity statistics and underlying fitness variance and mutation rate.
major comments (2)
- [Main derivation (following abstract description of surprisal decomposition)] The central derivation applies mean approximations to the decomposed surprisal rates to close the relation for the localization factor, yet no error bounds, fluctuation estimates, or convergence conditions are supplied for arbitrary landscapes. This is load-bearing for the generality claim, especially near the error threshold where delocalization is expected.
- [Application section (Hill-number combination)] No numerical validation or counter-example tests are presented to confirm that the mean closure remains accurate when fitness variance is high or when the population is partially delocalized. Without such checks the asserted applicability to arbitrary landscapes rests on an unverified assumption.
minor comments (2)
- [Notation and definitions] The precise definition of 'effective fitness variance' should be given an explicit equation label and computational formula so that readers can reproduce the localization factor from a given landscape.
- [Results/figures] A short table or figure comparing the predicted localization factor against direct simulation on at least two qualitatively different landscapes would greatly improve clarity and support the claims.
Simulated Author's Rebuttal
We thank the referee for their careful reading and constructive feedback on our manuscript. We agree that the validity of the mean approximations and their applicability require further clarification and support. We address each major comment below and will revise the manuscript accordingly to strengthen the presentation.
read point-by-point responses
-
Referee: The central derivation applies mean approximations to the decomposed surprisal rates to close the relation for the localization factor, yet no error bounds, fluctuation estimates, or convergence conditions are supplied for arbitrary landscapes. This is load-bearing for the generality claim, especially near the error threshold where delocalization is expected.
Authors: We acknowledge that the derivation relies on mean approximations without explicit error bounds or convergence criteria. This is a genuine limitation for the strongest generality claim. In the revised manuscript we will add a dedicated subsection on the approximation's regime of validity. This will include (i) a heuristic error estimate derived from the variance of the surprisal rates, (ii) references to existing fluctuation analyses in quasispecies theory, and (iii) a qualitative discussion of behavior near the error threshold, where the localization factor is expected to approach zero. We will also state the assumptions (large population size, separation of timescales) more explicitly. revision: yes
-
Referee: No numerical validation or counter-example tests are presented to confirm that the mean closure remains accurate when fitness variance is high or when the population is partially delocalized. Without such checks the asserted applicability to arbitrary landscapes rests on an unverified assumption.
Authors: We agree that numerical checks are necessary to support the claim of applicability. In the revised version we will include a new results subsection containing stochastic simulations on both single-peak and multi-peak landscapes. These will cover regimes of high fitness variance and mutation rates near the error threshold, comparing the predicted localization factor (from the Hill-number combination) against direct computation of the quasispecies distribution. We will report the relative error as a function of population size and landscape ruggedness. revision: yes
Circularity Check
No significant circularity; derivation is self-contained via mean-field approximations
full rationale
The paper derives a relation between combinations of Hill numbers and the localization factor (defined as effective fitness variance divided by squared mean mutation rate) by applying mean approximations to decomposed surprisal or stochastic entropy change rates under Eigen's model. The localization factor is constructed directly from the model's fitness and mutation parameters rather than being fitted to or redefined in terms of the Hill numbers. No load-bearing step reduces by construction to a self-definition, a fitted input renamed as prediction, or a self-citation chain. The mean approximations constitute an explicit modeling assumption whose validity bounds are a separate correctness question, not a circularity issue. The derivation therefore remains independent of its target outputs.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Mean approximations of decomposed surprisal or stochastic entropy change rates are valid across arbitrary fitness landscapes and mutation rates
Reference graph
Works this paper leans on
-
[1]
and define information as the Kullback-Leibler divergence betweenp ∗ and its equivalent constant-fitness quasispecies: IKL :=−S(p ∗)− NX i=0 p∗ i logb pµ i ,(39) wherep µ i denote the equilibrium frequencies of types sharing a replication rate (but with the same mutation probabilities that producedp ∗). This definition removes the confounding created by m...
-
[2]
M. Eigen, Selforganization of matter and the evolution of bio- logical macromolecules, Die Naturwissenschaften58, 465–523 (1971)
work page 1971
-
[3]
P. D. Sniegowski, P. J. Gerrish, T. Johnson, and A. Shaver, The evolution of mutation rates: separating causes from conse- quences, BioEssays22, 1057–1066 (2000)
work page 2000
-
[4]
R. Sanju ´an and P. Domingo-Calap, Mechanisms of viral mu- tation, Cellular and Molecular Life Sciences73, 4433–4448 (2016)
work page 2016
-
[5]
M. A. Nowak, What is a quasispecies?, Trends in Ecology & Evolution7, 118–121 (1992)
work page 1992
-
[6]
Domingo, Quasispecies theory in virology, Journal of Virol- ogy76, 463–465 (2002)
E. Domingo, Quasispecies theory in virology, Journal of Virol- ogy76, 463–465 (2002)
work page 2002
-
[7]
E. Domingo,Virus as Populations: Composition, Complex- ity, Quasispecies, Dynamics, and Biological Implications(Aca- demic Press, 2019)
work page 2019
-
[8]
J. F. Crow and M. Kimura,An Introduction to Population Ge- netics Theory(Harper and Row, New York, 1970)
work page 1970
-
[9]
P. Tarazona, Error thresholds for molecular quasispecies as phase transitions: From simple landscapes to spin-glass models, Physical Review A45, 6038–6050 (1992)
work page 1992
-
[10]
M. Eigen, Error catastrophe and antiviral strategy, Proceedings of the National Academy of Sciences99, 13374–13376 (2002)
work page 2002
-
[11]
I. Leuth ¨ausser, Statistical mechanics of Eigen’s evolution model, Journal of Statistical Physics48, 343–360 (1987)
work page 1987
-
[12]
S. Galluccio, Exact solution of the quasispecies model in a sharply peaked fitness landscape, Physical Review E56, 4526–4539 (1997)
work page 1997
-
[13]
D. B. Saakian and C.-K. Hu, Exact solution of the Eigen model with general fitness functions and degradation rates, Proceed- 12 ings of the National Academy of Sciences103, 4935–4939 (2006)
work page 2006
-
[14]
M. Nowak and P. Schuster, Error thresholds of replication in finite populations mutation frequencies and the onset of muller’s ratchet, Journal of Theoretical Biology137, 375–395 (1989)
work page 1989
-
[15]
D. Alves and J. F. Fontanari, Error threshold in finite popula- tions, Physical Review E57, 7008–7013 (1998)
work page 1998
-
[16]
P. R. A. Campos and J. F. Fontanari, Finite-size scaling of the error threshold transition in finite populations, Journal of Physics A: Mathematical and General32, L1–L7 (1999)
work page 1999
-
[17]
M. C. Boerlijst, S. Bonhoeffer, and M. A. Nowak, Viral quasi- species and recombination, Proceedings of the Royal Society of London. Series B: Biological Sciences263, 1577–1584 (1996)
work page 1996
-
[18]
M. N. Jacobi and M. Nordahl, Quasispecies and recombination, Theoretical Population Biology70, 479–485 (2006)
work page 2006
-
[19]
E. Tannenbaum and E. I. Shakhnovich, Error and repair catas- trophes: A two-dimensional phase diagram in the quasispecies model, Physical Review E69, 10.1103/physreve.69.011902 (2004)
-
[20]
E. Tannenbaum and E. I. Shakhnovich, Solution of the quasis- pecies model for an arbitrary gene network, Physical Review E 70, 10.1103/physreve.70.021903 (2004)
-
[22]
J. S. McCaskill, A localization threshold for macromolecular quasispecies from continuously distributed replication rates, The Journal of Chemical Physics80, 5194–5202 (1984)
work page 1984
-
[23]
B. Waclaw, Random matrices and localization in the quasis- pecies theory, Acta Physica Polonica B42, 1141 (2011)
work page 2011
-
[24]
E. Crosato, R. E. Spinney, and R. G. Morris, Stochastic mul- tistability of clonallike states in the Eigen model: a fidelity catastrophe (2025)
work page 2025
-
[25]
T. Wiehe, Model dependency of error thresholds: the role of fitness functions and contrasts between the finite and infinite sites models, Genetical Research69, 127–136 (1997)
work page 1997
-
[26]
E. Baake and H. Wagner, Mutation–selection models solved ex- actly with methods of statistical mechanics, Genetical Research 78, 93–117 (2001)
work page 2001
-
[27]
J. J. Bull, L. A. Meyers, and M. Lachmann, Quasispecies made simple, PLoS Computational Biology1, e61 (2005)
work page 2005
-
[28]
C. O. Wilke, Quasispecies theory in the context of population genetics, BMC Evolutionary Biology5, 10.1186/1471-2148-5- 44 (2005)
-
[29]
N. Takeuchi and P. Hogeweg, Error-threshold exists in fitness landscapes with lethal mutants, BMC Evolutionary Biology7, 15 (2007)
work page 2007
-
[30]
E. Domingo, D. Sabo, T. Taniguchi, and C. Weissmann, Nu- cleotide sequence heterogeneity of an RNA phage population, Cell13, 735–744 (1978)
work page 1978
-
[31]
S. Ito, Stochastic thermodynamic interpretation of informa- tion geometry, Physical Review Letters121, 10.1103/phys- revlett.121.030605 (2018)
-
[32]
S. Ito and A. Dechant, Stochastic time evolution, information geometry, and the cram ´er-rao bound, Physical Review X10, 10.1103/physrevx.10.021056 (2020)
-
[33]
S. B. Nicholson, L. P. Garc ´ıa-Pintos, A. del Campo, and J. R. Green, Time–information uncertainty relations in thermody- namics, Nature Physics16, 1211–1215 (2020)
work page 2020
- [34]
-
[35]
K. Adachi, R. Iritani, and R. Hamazaki, Universal constraint on nonlinear population dynamics, Communications Physics5, 10.1038/s42005-022-00912-4 (2022)
-
[36]
M. Hoshino, R. Nagayama, K. Yoshimura, J. F. Yamagishi, and S. Ito, Geometric speed limit for acceleration by natural selection in evolutionary processes, Physical Review Research 5, 10.1103/physrevresearch.5.023127 (2023)
-
[37]
L. P. Garc´ıa-Pintos, Limits on the evolutionary rates of biolog- ical traits, Scientific Reports14, 10.1038/s41598-024-61872-z (2024)
-
[38]
Quasilocalization under coupled mutation-selection dynamics
C. J. Palpal-latoc, Accompanying code for “Quasilocalization under coupled mutation-selection dynamics” (2026), GitHub repository
work page 2026
-
[39]
J. D. Hunter, Matplotlib: A 2d graphics environment, Comput- ing in Science & Engineering9, 90 (2007)
work page 2007
-
[40]
C. R. Harris, K. J. Millman, S. J. van der Walt, R. Gommers, P. Virtanen, D. Cournapeau, E. Wieser, J. Taylor, S. Berg, N. J. Smith, R. Kern, M. Picus, S. Hoyer, M. H. van Kerkwijk, M. Brett, A. Haldane, J. F. del R ´ıo, M. Wiebe, P. Peterson, P. G ´erard-Marchant, K. Sheppard, T. Reddy, W. Weckesser, H. Abbasi, C. Gohlke, and T. E. Oliphant, Array progra...
work page 2020
-
[41]
P. Virtanen, R. Gommers, T. E. Oliphant, M. Haber- land, T. Reddy, D. Cournapeau, E. Burovski, P. Peterson, W. Weckesser, J. Bright, S. J. van der Walt, M. Brett, J. Wilson, K. J. Millman, N. Mayorov, A. R. J. Nelson, E. Jones, R. Kern, E. Larson, C. J. Carey,˙I. Polat, Y. Feng, E. W. Moore, J. Vander- Plas, D. Laxalde, J. Perktold, R. Cimrman, I. Henriks...
work page 2020
-
[42]
pandas development team, pandas-dev/pandas: Pandas (2020)
T. pandas development team, pandas-dev/pandas: Pandas (2020)
work page 2020
-
[43]
Wes McKinney, Data Structures for Statistical Computing in Python, inProceedings of the 9th Python in Science Conference, edited by St´efan van der Walt and Jarrod Millman (2010) pp. 56 – 61
work page 2010
-
[44]
In this early stage, stochastic effects dominate and Eigen’s model therefore does not apply
Real viral populations start out with a very small population size after undergoing a population bottleneck at the start of an infection. In this early stage, stochastic effects dominate and Eigen’s model therefore does not apply. But unless the infection is cleared early on, the population size will eventually grow very large and the deterministic limit ...
-
[45]
1 isabsoluteandPN i=1 pi(t) =n tot(t) =constant
In Eigen’s original formulation, the total population size is constantso that the frequencyp i(t)in Eq. 1 isabsoluteandPN i=1 pi(t) =n tot(t) =constant. But Eq. 1 also holds for a population with atime-varyingtotal population size with the frequencyp i(t)beingrelativeand PN i=1 pi(t) = 1. Indeed, Eq. 1 is exact for the mean relative frequencies of the unc...
-
[46]
1 to model evolving phe- notypes or a more abstract population, biological or otherwise
Nothing however prevents applying Eq. 1 to model evolving phe- notypes or a more abstract population, biological or otherwise
- [47]
-
[48]
C. J. Thompson and J. L. McBride, On Eigen’s theory of the self- organization of matter and the evolution of biological macro- 13 molecules, Mathematical Biosciences21, 127–142 (1974)
work page 1974
-
[49]
Perron, Zur Theorie der Matrices, Mathematische Annalen 64, 248–263 (1907)
O. Perron, Zur Theorie der Matrices, Mathematische Annalen 64, 248–263 (1907)
work page 1907
-
[50]
G. Frobenius, ¨Uber Matrizen aus nicht negativen Elementen, Sitzungsberichte der Preußischen Akademie der Wissenschaften zu Berlin23, 456 (1912)
work page 1912
-
[51]
K. J. Arrow, A “dynamic” proof of the Frobenius–Perron theo- rem for Metzler matrices, inProbability, Statistics, and Mathe- matics(Elsevier, 1989) p. 17–26
work page 1989
-
[52]
Note also thatMis irreducible becauseW ′ is
Define the matrixM:=W ′ +kI, whereIis the identity matrix andkis some large enough constant such thatMis positive. Note also thatMis irreducible becauseW ′ is. Now,1is an eigenvector ofW′ and also ofM. The Perron-Frobenius theorem guarantees thatMhas a unique (up to a scalar multiple) positive eigenvector which corresponds to its largest eigenvalueλ M max...
-
[53]
G. E. Crooks, Entropy production fluctuation theorem and the nonequilibrium work relation for free energy differences, Phys. Rev. E60, 2721 (1999)
work page 1999
-
[54]
H. Qian, Entropy production and excess entropy in a nonequi- librium steady-state of single macromolecules, Phys. Rev. E65, 021111 (2002)
work page 2002
-
[55]
Entropy Production along a Stochastic Trajectory and an Integral Fluctuation Theorem,
U. Seifert, Entropy production along a stochastic trajectory and an integral fluctuation theorem, Physical Review Letters95, 10.1103/physrevlett.95.040602 (2005)
-
[56]
This is not unreasonable despite synonymous mutations that produce the same protein (and hence, supposedly the same fit- ness effect). In the case of SARS-CoV-2, for example, there is still variation in the fitness effects of synonymous mutations
-
[57]
and the site-specific mutation rates are even more variable [83]
-
[58]
That is,ϕincreases asx:= (x 1,
For a convex functiong, the sumϕ:= PN i=1 g(xi)is Schur- convex [84]. That is,ϕincreases asx:= (x 1, . . . , xN)becomes more localized. Then, the sums PN i=1 1/p∗ i and PN i=1(p∗ i )2, which have convex addends, both increase with increasing lo- calization, and as a result, so doesH(p ∗)
-
[59]
A. Stuart and K. Ord,Kendall’s advanced theory of statistics, distribution theory(John Wiley & Sons, 2010)
work page 2010
-
[60]
But consideringθ ∗ andθ † directly is equivalent
To be clear, they quantified the tendency of selection and mu- tation bounds to give tighter limits through theirP ∗(θ∗)and P †(θ†). But consideringθ ∗ andθ † directly is equivalent
-
[61]
Then,θ s =θ µ =⇒ (a−b)/c= (a−c)/b=⇒(b−c)(a−(b+c)) = 0
Leta=v info,b=v s, andc=v µ. Then,θ s =θ µ =⇒ (a−b)/c= (a−c)/b=⇒(b−c)(a−(b+c)) = 0. This has two solutions: eithera=b+corb=c. The former is a special case of the latter. After expandingv info and noting the identity √x+ √y= px+y+ 2 √xy, it can be written as Cov[ ˙σs,˙σµ] = p Var[ ˙σs]Var[ ˙σµ], where the covariance and variances are evaluated with respect...
-
[62]
Adami, Information theory in molecular biology, Physics of Life Reviews1, 3–22 (2004)
C. Adami, Information theory in molecular biology, Physics of Life Reviews1, 3–22 (2004)
work page 2004
-
[63]
Adami, What is complexity?, BioEssays24, 1085–1094 (2002)
C. Adami, What is complexity?, BioEssays24, 1085–1094 (2002)
work page 2002
-
[64]
C. Adami, What is information?, Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences374, 20150230 (2016)
work page 2016
-
[65]
C. Watkins, Selective breeding analysed as a communication channel: Channel capacity as a fundamental limit on adaptive complexity, in2008 10th International Symposium on Symbolic and Numeric Algorithms for Scientific Computing(2008) pp. 514–518
work page 2008
-
[66]
J. R. Peck and D. Waxman, Is life impossible? information, sex, and the origin of complex organisms, Evolution64, 3300–3309 (2010)
work page 2010
-
[67]
M. Hled ´ık, N. Barton, and G. Tkaˇcik, Accumulation and main- tenance of information in evolution, Proceedings of the National Academy of Sciences119, 10.1073/pnas.2123152119 (2022)
-
[68]
J. Gregori, C. Perales, F. Rodriguez-Frias, J. I. Esteban, J. Quer, and E. Domingo, Viral quasispecies complexity measures, Vi- rology493, 227–237 (2016)
work page 2016
-
[69]
E. Domingo and C. Perales, From quasispecies theory to viral quasispecies: How complexity has permeated virology, Mathe- matical Modelling of Natural Phenomena7, 105–122 (2012)
work page 2012
-
[70]
E. Domingo and P. Schuster, What is a quasispecies? historical origins and current scope, inQuasispecies: From Theory to Experimental Systems(Springer International Publishing, 2015) p. 1–22
work page 2015
-
[71]
J. K. Pfeiffer and K. Kirkegaard, Increased fidelity reduces po- liovirus fitness and virulence under selective pressure in mice, PLoS Pathogens1, e11 (2005)
work page 2005
-
[72]
M. Vignuzzi, J. K. Stone, J. J. Arnold, C. E. Cameron, and R. Andino, Quasispecies diversity determines pathogenesis through cooperative interactions in a viral population, Nature 439, 344–348 (2005)
work page 2005
- [73]
-
[74]
S. Ojosnegros, R. Agudo, M. Sierra, C. Briones, S. Sierra, C. Gonz´alez-L´opez, E. Domingo, and J. Cristina, Topology of evolving, mutagenized viral populations: quasispecies expan- sion, compression, and operation of negative selection, BMC Evolutionary Biology8, 10.1186/1471-2148-8-207 (2008)
-
[75]
C. Perales, M. Henry, E. Domingo, S. Wain-Hobson, and J.- P. Vartanian, Lethal mutagenesis of foot-and-mouth disease virus involves shifts in sequence space, Journal of Virology 85, 12227–12240 (2011)
work page 2011
-
[76]
M. O. Hill, Diversity and evenness: A unifying notation and its consequences, Ecology54, 427–432 (1973)
work page 1973
-
[77]
A. Chao, C.-H. Chiu, and L. Jost, Unifying species diversity, phylogenetic diversity, functional diversity, and related similar- ity and differentiation measures through hill numbers, Annual Review of Ecology, Evolution, and Systematics45, 297–324 (2014)
work page 2014
-
[78]
J. Gregori, M. Salicr´ u, E. Domingo, A. Sanchez, J. I. Esteban, F. Rodr´ıguez-Fr´ıas, and J. Quer, Inference with viral quasis- pecies diversity indices: clonal and NGS approaches, Bioinfor- matics30, 1104–1111 (2014)
work page 2014
-
[79]
S. Posada-Cespedes, D. Seifert, and N. Beerenwinkel, Recent advances in inferring viral diversity from high-throughput se- quencing data, Virus Research239, 17–32 (2017)
work page 2017
-
[80]
S. Knyazev, L. Hughes, P. Skums, and A. Zelikovsky, Epidemio- logical data analysis of viral quasispecies in the next-generation sequencing era, Briefings in Bioinformatics22, 96–108 (2020)
work page 2020
-
[81]
L. Fuhrmann, B. Langer, I. Topolsky, and N. Beerenwinkel, VILOCA: sequencing quality-aware viral haplotype reconstruc- tion and mutation calling for short-read and long-read data, NAR Genomics and Bioinformatics6, 10.1093/nargab/lqae152 (2024)
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.