Recognition: 2 theorem links
· Lean TheoremMSGL-Transformer: A Multi-Scale Global-Local Transformer for Rodent Social Behavior Recognition
Pith reviewed 2026-05-10 17:50 UTC · model grok-4.3
The pith
A multi-scale global-local transformer recognizes rodent social behaviors from pose sequences more accurately than prior models.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
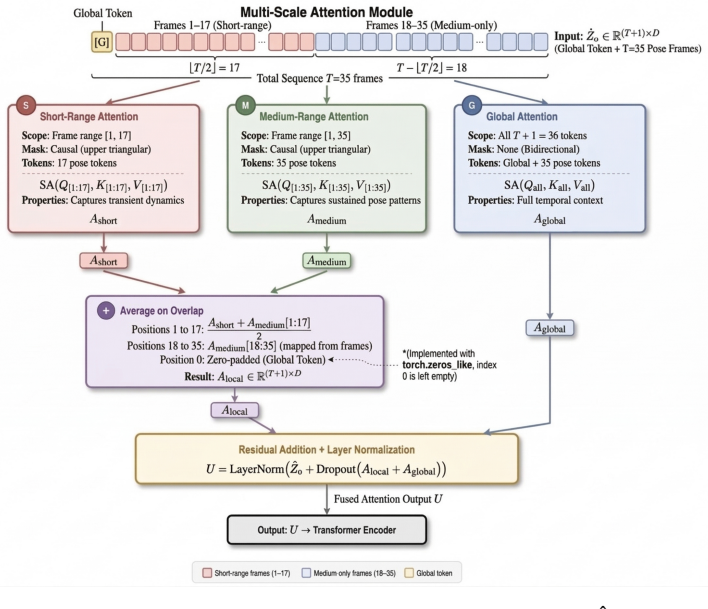
MSGL-Transformer uses a lightweight transformer encoder whose multi-scale attention consists of parallel short-range, medium-range, and global branches that explicitly capture motion dynamics at different temporal scales, combined with a Behavior-Aware Modulation block that adjusts temporal embeddings to highlight behavior-relevant features before attention is applied.
What carries the argument
Multi-scale attention mechanism formed by three parallel branches (short-range, medium-range, global) plus the Behavior-Aware Modulation (BAM) block that modulates embeddings in the style of squeeze-and-excitation networks.
If this is right
- Outperforms TCN, LSTM, and Bi-LSTM baselines on the RatSI dataset, reaching 75.4 percent mean accuracy across nine cross-validation splits.
- Achieves 87.1 percent accuracy and 0.8745 F1 on CalMS21, a 10.7 percent gain over HSTWFormer while also beating ST-GCN, MS-G3D, CTR-GCN, and STGAT.
- The identical architecture works on both five-class and four-class problems after changing only input dimensionality and number of output classes.
- Explicit separation of attention scales makes the contribution of each temporal range directly observable.
Where Pith is reading between the lines
- Success would make large-scale automated analysis of rodent social behavior practical for labs that currently rely on manual scoring.
- The design could be tested on pose data from other species or on behaviors that involve more than two animals.
- Future work could measure how much performance depends on the quality of the upstream pose tracker by injecting controlled tracking errors.
Load-bearing premise
The 12-dimensional or 28-dimensional pose keypoints supplied as input are accurate and complete representations of the animals' movements.
What would settle it
Apply the trained model to the same videos but replace the clean keypoints with versions that contain realistic tracking noise or missing joints and measure whether accuracy falls substantially below the reported 75.4 percent and 87.1 percent figures.
Figures









read the original abstract
Recognition of rodent behavior is important for understanding neural and behavioral mechanisms. Traditional manual scoring is time-consuming and prone to human error. We propose MSGL-Transformer, a Multi-Scale Global-Local Transformer for recognizing rodent social behaviors from pose-based temporal sequences. The model employs a lightweight transformer encoder with multi-scale attention to capture motion dynamics across different temporal scales. The architecture integrates parallel short-range, medium-range, and global attention branches to explicitly capture behavior dynamics at multiple temporal scales. We also introduce a Behavior-Aware Modulation (BAM) block, inspired by SE-Networks, which modulates temporal embeddings to emphasize behavior-relevant features prior to attention. We evaluate on two datasets: RatSI (5 behavior classes, 12D pose inputs) and CalMS21 (4 behavior classes, 28D pose inputs). On RatSI, MSGL-Transformer achieves 75.4% mean accuracy and F1-score of 0.745 across nine cross-validation splits, outperforming TCN, LSTM, and Bi-LSTM. On CalMS21, it achieves 87.1% accuracy and F1-score of 0.8745, a +10.7% improvement over HSTWFormer, and outperforms ST-GCN, MS-G3D, CTR-GCN, and STGAT. The same architecture generalizes across both datasets with only input dimensionality and number of classes adjusted.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript presents MSGL-Transformer, a Multi-Scale Global-Local Transformer for rodent social behavior recognition from pose sequences. It features a lightweight transformer with parallel short-, medium-, and global-range attention branches, along with a Behavior-Aware Modulation (BAM) block to emphasize relevant features. Evaluations on the RatSI (5 classes, 12D pose) and CalMS21 (4 classes, 28D pose) datasets report mean accuracies of 75.4% and 87.1%, respectively, with F1 scores of 0.745 and 0.8745, outperforming baselines including TCN, LSTM, Bi-LSTM, HSTWFormer, ST-GCN, MS-G3D, CTR-GCN, and STGAT. The architecture is claimed to generalize across datasets with only adjustments to input dimensions and class counts.
Significance. If the performance claims are substantiated through additional verification, the work offers a promising direction for automated analysis of rodent social behaviors, which is valuable for neuroscience and behavioral studies. The multi-scale attention mechanism addresses the temporal variability in behaviors, and the consistent architecture across two datasets demonstrates potential for broader applicability. The use of public datasets and direct comparisons to published baselines is a strength.
major comments (3)
- [Experimental Evaluation] The reported mean accuracy of 75.4% on RatSI across nine cross-validation splits and 87.1% on CalMS21 lack error bars, standard deviations, or statistical significance tests. This makes it challenging to determine whether the improvements over baselines such as TCN, LSTM, and HSTWFormer are statistically meaningful.
- [Method and Experiments] No ablation studies are provided for the multi-scale attention branches (short-range, medium-range, global) or the Behavior-Aware Modulation (BAM) block. Since these are the core innovations, their individual contributions to the reported F1 scores (0.745 on RatSI, 0.8745 on CalMS21) cannot be verified, undermining the central architectural claims.
- [Introduction and Evaluation] The model relies on 12D and 28D pose keypoints without any experiments testing sensitivity to tracking errors, occlusions, or missing joints. Given that rodent pose estimation is prone to such issues and the architecture lacks explicit noise-handling mechanisms, the generalization claim across datasets may not hold in practical, noisy conditions.
minor comments (2)
- [Abstract] The abstract states 'nine cross-validation splits' for RatSI but does not specify the split strategy or dataset size, which would aid reproducibility.
- [Method] Additional details on the exact integration of the BAM block with the attention branches would improve clarity of the method description.
Simulated Author's Rebuttal
We thank the referee for the constructive comments on our manuscript. We address each major comment point by point below and will incorporate revisions where they strengthen the work.
read point-by-point responses
-
Referee: The reported mean accuracy of 75.4% on RatSI across nine cross-validation splits and 87.1% on CalMS21 lack error bars, standard deviations, or statistical significance tests. This makes it challenging to determine whether the improvements over baselines such as TCN, LSTM, and HSTWFormer are statistically meaningful.
Authors: We agree that reporting variability and statistical tests would improve the evaluation section. The means are averaged over nine cross-validation splits on RatSI, yet standard deviations and error bars were omitted. In the revised manuscript we will add standard deviations for all metrics, include error bars in the tables, and perform paired statistical tests (e.g., Wilcoxon signed-rank) against the baselines to confirm the significance of the reported gains. revision: yes
-
Referee: No ablation studies are provided for the multi-scale attention branches (short-range, medium-range, global) or the Behavior-Aware Modulation (BAM) block. Since these are the core innovations, their individual contributions to the reported F1 scores (0.745 on RatSI, 0.8745 on CalMS21) cannot be verified, undermining the central architectural claims.
Authors: We acknowledge that the absence of ablation studies leaves the contribution of each proposed component unquantified. The original submission presented the full model and its overall results but did not isolate the branches or the BAM block. We will conduct the necessary ablation experiments for the revised version, reporting accuracy and F1 scores on both datasets when each attention branch and the BAM block are removed individually. revision: yes
-
Referee: The model relies on 12D and 28D pose keypoints without any experiments testing sensitivity to tracking errors, occlusions, or missing joints. Given that rodent pose estimation is prone to such issues and the architecture lacks explicit noise-handling mechanisms, the generalization claim across datasets may not hold in practical, noisy conditions.
Authors: This is a fair observation about real-world robustness. Although the architecture generalizes across the two datasets with different input dimensions, we did not evaluate performance under simulated pose-estimation artifacts. In the revision we will add controlled experiments that introduce missing joints, occlusions, and additive noise to the keypoint sequences and report the resulting performance drops on both datasets, together with a short discussion of possible noise-robust extensions. revision: yes
Circularity Check
No circularity: empirical ML evaluation on public datasets with independent baselines
full rationale
The paper describes a transformer architecture (multi-scale attention branches + BAM block) and reports accuracies/F1 scores obtained by training and evaluating on fixed public datasets (RatSI, CalMS21) under standard cross-validation. No equations, predictions, or uniqueness claims reduce the reported results to fitted constants, self-citations, or input redefinitions by construction. The architecture choices are presented as design decisions, not derived from prior self-work that would force the outcomes. Performance gains are measured against published external baselines, making the central claims self-contained and falsifiable outside any internal loop.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Pose keypoints provide a sufficient and low-noise representation of rodent social behavior
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
The model employs a lightweight transformer encoder with multi-scale attention... parallel short-range, medium-range, and global attention branches... Behavior-Aware Modulation (BAM) block
-
IndisputableMonolith/Foundation/ArithmeticFromLogic.leanLogicNat ≃ Nat recovery unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
same architecture generalizes across both datasets with only input dimensionality and number of classes adjusted
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
Animal use in neurobiological research.Neuroscience433, 1–10 (2020)
˙Zakowski, W. Animal use in neurobiological research.Neuroscience433, 1–10 (2020)
2020
-
[2]
& Youn, J
Ellenbroek, B. & Youn, J. Rodent models in neuroscience research: is it a rat race?Dis. models & mechanisms9, 1079–1087 (2016)
2016
-
[3]
Bryda, E. C. The mighty mouse: the impact of rodents on advances in biomedical research.Mo. medicine110, 207 (2013)
2013
-
[4]
Neuroanatomical substrates of rodent social behavior: the medial prefrontal cortex and its projection patterns.Front
Ko, J. Neuroanatomical substrates of rodent social behavior: the medial prefrontal cortex and its projection patterns.Front. neural circuits11, 41 (2017)
2017
-
[5]
Pharmacol.14, 1329424 (2024)
Popik, P.et al.Effects of ketamine on rat social behavior as analyzed by deeplabcut and simba deep learning algorithms.Front. Pharmacol.14, 1329424 (2024)
2024
-
[6]
& Marklund, N
Hånell, A. & Marklund, N. Structured evaluation of rodent behavioral tests used in drug discovery research.Front. behavioral neuroscience8, 252 (2014)
2014
-
[7]
A., Afzal, A., Warraich, Z
Desland, F. A., Afzal, A., Warraich, Z. & Mocco, J. Manual versus automated rodent behavioral assessment: comparing efficacy and ease of bederson and garcia neurological deficit scores to an open field video-tracking system.J. central nervous system disease6, JCNSD–S13194 (2014)
2014
-
[8]
learning memory165, 106780 (2019)
Gulinello, M.et al.Rigor and reproducibility in rodent behavioral research.Neurobiol. learning memory165, 106780 (2019)
2019
-
[9]
& Aguiar, P
Gerós, A., Magalhães, A. & Aguiar, P. Improved 3d tracking and automated classification of rodents’ behavioral activity using depth-sensing cameras.Behav. research methods52, 2156–2167 (2020). 11.V on Ziegler, L., Sturman, O. & Bohacek, J. Big behavior: challenges and opportunities in a new era of deep behavior profiling.Neuropsychopharmacology46, 33–44 (2021)
2020
-
[10]
S.et al.Beyond observation: Deep learning for animal behavior and ecological conservation
Saoud, L. S.et al.Beyond observation: Deep learning for animal behavior and ecological conservation. Ecol. Informatics102893 (2024)
2024
-
[11]
P.et al.Deepethogram, a machine learning pipeline for supervised behavior classification from raw pixels.elife10, e63377 (2021)
Bohnslav, J. P.et al.Deepethogram, a machine learning pipeline for supervised behavior classification from raw pixels.elife10, e63377 (2021)
2021
-
[12]
Segalin, C.et al.The mouse action recognition system (mars) software pipeline for automated analysis of social behaviors in mice.Elife10, e63720 (2021)
2021
-
[13]
Y .et al.Using deep learning to study emotional behavior in rodent models.Front
Kuo, J. Y .et al.Using deep learning to study emotional behavior in rodent models.Front. Behav. Neurosci.16, 1044492 (2022)
2022
-
[14]
A., Noldus, L
Van Dam, E. A., Noldus, L. P. & Van Gerven, M. A. Disentangling rodent behaviors to improve automated behavior recognition.Front. Neurosci.17, 1198209 (2023). 23/25
2023
-
[15]
& Stefanini, C
Fazzari, E., Romano, D., Falchi, F. & Stefanini, C. Animal behavior analysis methods using deep learning: A survey.Expert. Syst. With Appl.128330 (2025)
2025
-
[16]
neuroscience methods300, 166–172 (2018)
Lorbach, M.et al.Learning to recognize rat social behavior: Novel dataset and cross-dataset application.J. neuroscience methods300, 166–172 (2018)
2018
-
[17]
Fish, E., Weinbren, J. & Gilbert, A. Two-stream transformer architecture for long video understanding. arXiv preprint arXiv:2208.01753(2022). 20.Vaswani, A.et al.Attention is all you need.Adv. neural information processing systems30(2017)
-
[18]
I., Caragea, D
Sharif, M. I., Caragea, D. & Iqbal, A. Rodent social behavior recognition using a global context-aware vision transformer network.AI6, 264 (2025)
2025
-
[19]
InProceedings of the IEEE/CVF international conference on computer vision, 10012–10022 (2021)
Liu, Z.et al.Swin transformer: Hierarchical vision transformer using shifted windows. InProceedings of the IEEE/CVF international conference on computer vision, 10012–10022 (2021)
2021
-
[20]
InProceedings of the IEEE/CVF international conference on computer vision, 6836–6846 (2021)
Arnab, A.et al.Vivit: A video vision transformer. InProceedings of the IEEE/CVF international conference on computer vision, 6836–6846 (2021)
2021
-
[21]
J.et al.The multi-agent behavior dataset: Mouse dyadic social interactions.Adv
Sun, J. J.et al.The multi-agent behavior dataset: Mouse dyadic social interactions.Adv. neural information processing systems2021, 1 (2021)
2021
-
[22]
& Sun, G
Hu, J., Shen, L. & Sun, G. Squeeze-and-excitation networks. InProceedings of the IEEE conference on computer vision and pattern recognition, 7132–7141 (2018)
2018
-
[23]
& Duan, F
Ru, Z. & Duan, F. Hierarchical spatial–temporal window transformer for pose-based rodent behavior recognition.IEEE Transactions on Instrumentation Meas.73, 1–14 (2024)
2024
-
[24]
& Lin, D
Yan, S., Xiong, Y . & Lin, D. Spatial temporal graph convolutional networks for skeleton-based action recognition. InProceedings of the AAAI Conference on Artificial Intelligence(2018)
2018
-
[25]
& Ouyang, W
Liu, Z., Zhang, H., Chen, Z., Wang, Z. & Ouyang, W. Disentangling and unifying graph convolutions for skeleton-based action recognition. InProceedings of the IEEE/CVF Conference on Computer Vision and Pattern Recognition, 143–152 (2020)
2020
-
[26]
InProceedings of the IEEE/CVF International Conference on Computer Vision, 13359– 13368 (2021)
Chen, Y .et al.Channel-wise topology refinement graph convolution for skeleton-based action recognition. InProceedings of the IEEE/CVF International Conference on Computer Vision, 13359– 13368 (2021)
2021
-
[27]
& Van Gool, L
Huang, Z., Wan, C., Probst, T. & Van Gool, L. Spatial-temporal graph attention networks for skeleton-based action recognition.J. Electron. Imaging29(2020)
2020
-
[28]
Anderson, D. J. & Perona, P. Toward a science of computational ethology.Neuron84, 18–31 (2014)
2014
-
[29]
Methods9, 410–417 (2012)
de Chaumont, F.et al.Computerized video analysis of social interactions in mice.Nat. Methods9, 410–417 (2012). 33.Yu, X. e. a. Automated home-cage behavioral phenotyping of mice.Nat. Commun.1, 1–9 (2009)
2012
-
[30]
Giancardo, L.et al.Automatic visual tracking and social behaviour analysis with multiple mice.PloS one8, e74557 (2013)
2013
-
[31]
P., Dollár, P., Lin, D., Anderson, D
Burgos-Artizzu, X. P., Dollár, P., Lin, D., Anderson, D. J. & Bhatt Perona, P. Social behavior recogni- tion in continuous video. In2012 IEEE Conference on Computer Vision and Pattern Recognition, 1322–1329 (IEEE, 2012)
2012
-
[32]
Mathis, M. W. & Mathis, A. Deep learning tools for the measurement of animal behavior in neuroscience.Curr. opinion neurobiology60, 1–11 (2020). 24/25
2020
-
[33]
& Haffner, P
LeCun, Y ., Bottou, L., Bengio, Y . & Haffner, P. Gradient-based learning applied to document recognition.Proc. IEEE86, 2278–2324 (1998). 38.Hochreiter, S. & Schmidhuber, J. Long short-term memory.Neural Comput.9, 1735–1780 (1997)
1998
-
[34]
Reports15, 4982, DOI: 10.1038/s41598-025-87752-8 (2025)
Chen, D., Chen, M., Wu, P.et al.Two-stream spatio-temporal GCN-transformer networks for skeleton-based action recognition.Sci. Reports15, 4982, DOI: 10.1038/s41598-025-87752-8 (2025)
-
[35]
Mlost, J., Dawli, R., Liu, X., Costa, A. R. & Dorocic, I. P. Evaluation of unsupervised learning algorithms for the classification of behavior from pose estimation data.Patterns6(2025)
2025
-
[36]
R.et al.Simple behavioral analysis (simba)–an open source toolkit for computer classification of complex social behaviors in experimental animals.BioRxiv2020–04 (2020)
Nilsson, S. R.et al.Simple behavioral analysis (simba)–an open source toolkit for computer classification of complex social behaviors in experimental animals.BioRxiv2020–04 (2020)
2020
-
[37]
Luxem, K.et al.Identifying behavioral structure from deep variational embeddings of animal motion. Commun. Biol.5, 1267 (2022)
2022
-
[38]
An Image is Worth 16x16 Words: Transformers for Image Recognition at Scale
Dosovitskiy, A.et al.An image is worth 16x16 words: Transformers for image recognition at scale. arXiv preprint arXiv:2010.11929(2020)
work page internal anchor Pith review Pith/arXiv arXiv 2010
-
[39]
InThe Fourteenth International Conference on Learning Representations(2026)
Wang, Y .et al.Animal behavioral analysis and neural encoding with transformer-based self-supervised pretraining. InThe Fourteenth International Conference on Learning Representations(2026)
2026
-
[40]
machine Learn
Pedregosa, F.et al.Scikit-learn: Machine learning in python.J. machine Learn. research12, 2825–2830 (2011)
2011
-
[41]
neural information processing systems32(2019)
Paszke, A.et al.Pytorch: An imperative style, high-performance deep learning library.Adv. neural information processing systems32(2019). 47.Fan, H.et al.Multiscale vision transformers. InICCV(2021)
2019
-
[42]
& Liu, H
Chen, Z., Li, S., Yang, B., Li, Q. & Liu, H. Multi-scale spatial temporal graph convolutional network for skeleton-based action recognition.AAAI(2021)
2021
-
[43]
Bai, S., Kolter, J. Z. & Koltun, V . An empirical evaluation of generic convolutional and recurrent networks for sequence modeling. arxiv.arXiv preprint arXiv:1803.0127110(2018)
work page internal anchor Pith review arXiv 2018
-
[44]
& Paliwal, K
Schuster, M. & Paliwal, K. K. Bidirectional recurrent neural networks.IEEE Transactions on Signal Process.(1997). 25/25
1997
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.