Recognition: unknown
Fixation probabilities for multi-allele Moran dynamics with weak selection
Pith reviewed 2026-05-10 15:55 UTC · model grok-4.3
The pith
Fixation probabilities for multiple competing alleles under weak selection expand systematically around the neutral solution.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
We develop a perturbative framework to compute fixation probabilities in multi-allele Moran processes under weak selection. Exploiting the general structure of the backward Fokker-Planck operator in this regime, we show that fixation probabilities admit a systematic expansion around their neutral solution for M competing alleles and arbitrary fitness functions.
What carries the argument
The backward Fokker-Planck operator of the multi-allele Moran process, whose weak-selection structure permits a perturbative expansion of fixation probabilities around the neutral solution.
If this is right
- Fixation probabilities become analytically accessible for a simple three-allele model with constant fitness.
- The expansion applies to coordination games where allele fitness increases with its own frequency.
- The method handles clonal interference between mutualistic alleles.
- Analytical results for fixation extend beyond pairwise interactions to general multi-strategy dynamics.
Where Pith is reading between the lines
- The same perturbative structure might apply to other birth-death processes with weak selection if their transition rates admit a similar expansion.
- Higher-order terms in the series could be computed explicitly to improve accuracy for moderate selection strengths.
- The framework could be tested on frequency-dependent fitness functions beyond the three examples given.
Load-bearing premise
The backward Fokker-Planck operator in the weak-selection regime has a structure that permits a systematic perturbative expansion of fixation probabilities without breakdown or need for resummation.
What would settle it
Direct numerical simulation of the Moran process for three alleles with a chosen fitness function, followed by comparison of the computed fixation probabilities against the perturbative series truncated at low orders.
Figures


read the original abstract
Fixation probabilities are essential for characterizing stochastic evolutionary dynamics, but analytical results remain limited mainly to systems with two competing types. We develop a perturbative framework to compute fixation probabilities in multi-allele Moran processes under weak selection. Exploiting the general structure of the backward Fokker-Planck operator in this regime, we show that fixation probabilities admit a systematic expansion around their neutral solution. We first introduce the framework in a general case with $M$ competing alleles and arbitrary fitness functions, and then apply it to three biologically motivated examples: a simple model of three competing alleles with a constant fitness function, a coordination game in which allele fitness increases with its frequency in the population, and a model of clonal interference between mutualistic alleles. These results extend the analytical understanding of fixation probabilities beyond pairwise interactions, establishing a framework for investigating multi-strategy stochastic evolutionary dynamics.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript develops a perturbative framework to compute fixation probabilities for multi-allele Moran processes under weak selection. It expands the fixation probability as a power series in selection strength around the neutral solution by exploiting the structure of the backward Fokker-Planck operator, first presenting the general case for M alleles with arbitrary fitness functions and then applying the method to three examples: constant fitness for three alleles, a coordination game with frequency-dependent fitness, and clonal interference between mutualistic alleles.
Significance. If the expansion is rigorously justified, the work would extend analytical results on fixation probabilities beyond the two-allele limit and supply a systematic tool for multi-strategy stochastic evolutionary dynamics. The approach builds on the neutral operator's kernel properties to generate recursive corrections, which is a strength when the solvability conditions hold.
major comments (2)
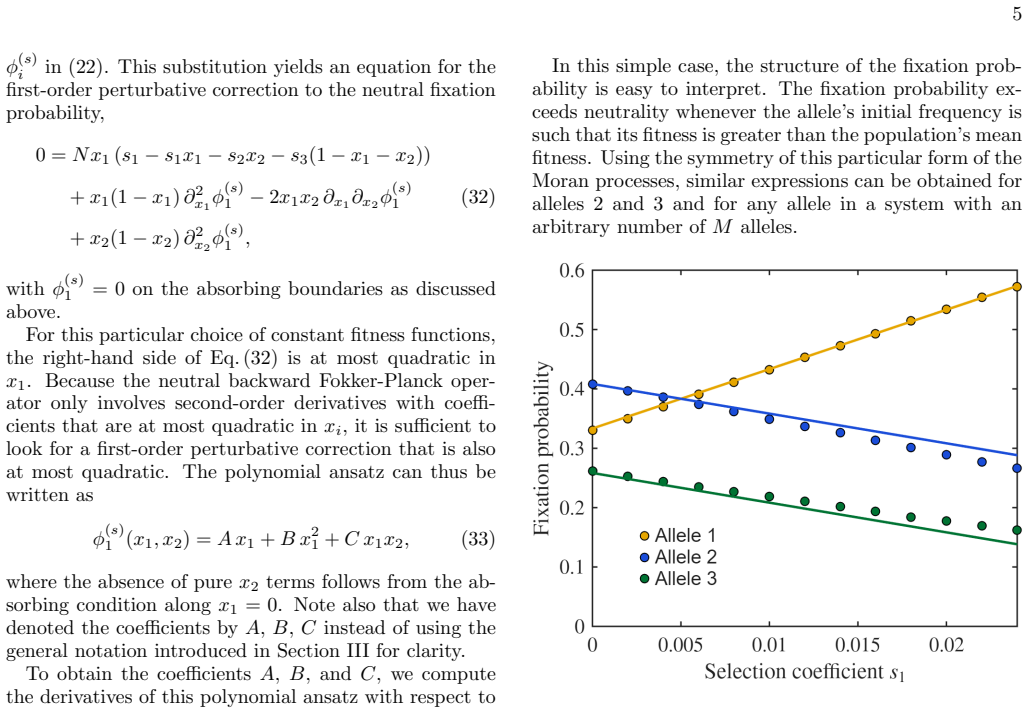
- [§2 (general framework)] §2 (general framework): The central claim that the expansion is systematic for arbitrary fitness functions rests on the assumption that solvability conditions for the neutral backward operator L_0 are automatically satisfied at each order. The source term at order n must be orthogonal to the adjoint kernel (constants or neutral fixation probabilities on the simplex). The manuscript does not derive or verify this orthogonality for general fitness; it holds in the three examples by symmetry or construction but is not guaranteed a priori for arbitrary fitness, risking non-existence of solutions or secular terms.
- [Abstract and §3–5 (examples)] Abstract and §3–5 (examples): The three applications are presented as support for the general framework, but without explicit checks (e.g., verification that the source terms satisfy the Fredholm alternative at each perturbative order or comparison to exact neutral limits), it is unclear whether the hierarchy closes without resummation. This weakens the assertion that the method works for arbitrary fitness.
minor comments (2)
- [Abstract] Abstract: The phrase 'arbitrary fitness functions' is used without qualification; a brief caveat that the expansion assumes solvability conditions would improve precision.
- [Notation] Notation: Define the selection-strength parameter and the fitness perturbation operator consistently between the general derivation and the example sections to avoid ambiguity in the perturbative hierarchy.
Simulated Author's Rebuttal
We thank the referee for the careful and constructive review of our manuscript. The comments raise important points about the rigor of the general perturbative framework and the supporting examples. We address each major comment below and indicate the revisions we will make to strengthen the presentation.
read point-by-point responses
-
Referee: [§2 (general framework)] The central claim that the expansion is systematic for arbitrary fitness functions rests on the assumption that solvability conditions for the neutral backward operator L_0 are automatically satisfied at each order. The source term at order n must be orthogonal to the adjoint kernel (constants or neutral fixation probabilities on the simplex). The manuscript does not derive or verify this orthogonality for general fitness; it holds in the three examples by symmetry or construction but is not guaranteed a priori for arbitrary fitness, risking non-existence of solutions or secular terms.
Authors: We thank the referee for identifying this key aspect of the justification. The manuscript presents the general framework in §2 by exploiting the kernel structure of the neutral backward operator, but it is correct that an explicit derivation of the orthogonality (Fredholm alternative) condition for arbitrary fitness functions is not provided. While the construction ensures that fitness perturbations vanish in the neutral limit, a general proof that source terms remain orthogonal to the adjoint kernel at every order was omitted. We will revise §2 to include this derivation, showing that the inner product of each source term with the neutral fixation probabilities integrates to zero over the simplex by the properties of the Moran transition rates and the definition of the selection perturbation. This addition will confirm that solutions exist order by order without secular terms for arbitrary fitness. revision: yes
-
Referee: [Abstract and §3–5 (examples)] The three applications are presented as support for the general framework, but without explicit checks (e.g., verification that the source terms satisfy the Fredholm alternative at each perturbative order or comparison to exact neutral limits), it is unclear whether the hierarchy closes without resummation. This weakens the assertion that the method works for arbitrary fitness.
Authors: We agree that the examples would benefit from explicit verification to demonstrate that the perturbative hierarchy is well-defined. In the revised manuscript, we will augment §§3–5 with direct checks for each example: (i) confirmation that the source terms at each order are orthogonal to the adjoint kernel, (ii) explicit reduction to the exact neutral fixation probabilities when the selection coefficient is set to zero, and (iii) verification that no resummation is required because the solvability condition holds at every finite order. These additions will be concise and will reinforce that the framework applies beyond the specific cases shown. revision: yes
Circularity Check
No circularity: expansion follows from standard backward Fokker-Planck structure
full rationale
The derivation begins from the established backward Fokker-Planck operator for the multi-allele Moran process under weak selection, a standard stochastic-process tool independent of the paper. The perturbative hierarchy is constructed order-by-order by applying this operator to successive corrections, with the neutral solution serving as the zeroth-order input. No step defines the target fixation probabilities in terms of themselves, renames a fitted quantity as a prediction, or relies on a load-bearing self-citation whose content is unverified outside the present work. The three examples are downstream applications, not inputs that force the general result. The framework is therefore self-contained against external mathematical benchmarks.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption The Moran process dynamics can be approximated by a backward Fokker-Planck equation under weak selection.
- standard math Neutral (zero-selection) fixation probabilities are known exactly and form a valid base for perturbation.
Reference graph
Works this paper leans on
-
[1]
Computing the relevant derivatives of this ansatz with respect to allele frequencies, and substituting them in Eq
This equation suggests a multivariate polynomial ansatz of the form ϕ(s) 1 =N Ax1 +Bx 2 1 +Cx 3 1 +Dx 1x2 +Ex 2 1x2 +F x 1x2 2 , (40) where, again, the absence of purex 2 terms follows from the absorbing condition alongx 1 = 0. Computing the relevant derivatives of this ansatz with respect to allele frequencies, and substituting them in Eq. (39), we obtai...
-
[2]
Fixation probability perturbationϕ (s) 1 =ϕ 1 −x 1 for allele 1 in the three-allele Moran process under frequency- dependent selection
The multivariate polynomial ansatz has the same 7 Figure 3. Fixation probability perturbationϕ (s) 1 =ϕ 1 −x 1 for allele 1 in the three-allele Moran process under frequency- dependent selection. The color map represents the analytical solution over the simplex, while streamlines indicate the direction of maximal increase of the perturbation. (a) In the o...
2024
-
[3]
RMG received partial support from the S˜ ao Paulo Research Foundation (FAPESP) through grant ICTP- SAIFR 2021/14335-0
2021
-
[4]
1978;40(1–2):145–156.doi:10.1016/0025-5564(78)90077- 9
Taylor PD, Jonker LB.Evolutionarily stable strate- gies and game dynamics.Mathematical Biosciences. 1978;40(1–2):145–156.doi:10.1016/0025-5564(78)90077- 9
-
[5]
2009;2:25–61.doi:10.1002/9783527620304.ch2
Traulsen A, Hauert C.Stochastic evolutionary game dy- namics.Reviews of Nonlinear Dynamics and Complexity. 2009;2:25–61.doi:10.1002/9783527620304.ch2
-
[6]
G. W. A. Constable, A. J. McKane, T. Rogers, and C. E. Tarnita, Demographic noise can reverse the direction of deterministic selection,Proc. Natl. Acad. Sci. USA113, E4745–E4754 (2016). 9
2016
-
[7]
Nowak MA.Evolutionary Dynamics: Exploring the Equa- tions of Life.Cambridge: Harvard University Press; 2006
2006
-
[8]
The Japanese Journal of Ge- netics; 1991
Kimura M.The neutral theory of molecular evolution: a review of recent evidence. The Japanese Journal of Ge- netics; 1991
1991
-
[9]
2018;2(7):1155– 1160.doi:10.1038/s41559-018-0569-4
Gore AV, Tomins KA, Iben J, Ma L, Castranova D, Davis AE, Parkhurst A, Jeffery WR, Weinstein BM.An epigenetic mechanism for cavefish eye degen- eration.Nature Ecology & Evolution. 2018;2(7):1155– 1160.doi:10.1038/s41559-018-0569-4
-
[10]
2016;113(17):4765– 4770.doi:10.1073/pnas.1522931113
Wright NA, Steadman DW, Witt CC.Predictable evolution toward flightlessness in volant is- land birds.Proceedings of the National Academy of Sciences USA. 2016;113(17):4765– 4770.doi:10.1073/pnas.1522931113
-
[11]
1958;54(1):60–71.doi:10.1017/S0305004100033193
Moran PAP.Random processes in genetics.Mathematical Proceedings of the Cambridge Philosophical Society. 1958;54(1):60–71.doi:10.1017/S0305004100033193
-
[12]
2021;39:100347.doi:10.1016/j.cosrev.2020.100347
D´ ıaz J, Goldberg LA, Mertzios GB, Richerby D, Serna M, Spirakis PG.A survey of the modified Moran process and evolutionary graph theory.Computer Science Review. 2021;39:100347.doi:10.1016/j.cosrev.2020.100347
-
[13]
D. M. McCandlish, C. L. Epstein, and J. B. Plotkin, Formal properties of the probability of fixation: Identities, inequalities and approximations,Theor. Popul. Biol.99, 98–113 (2015)
2015
-
[14]
2009;80(1):011909.doi:10.1103/PhysRevE.80.011909
Altrock PM, Traulsen A.Deterministic evolutionary game dynamics in finite populations.Physical Review E. 2009;80(1):011909.doi:10.1103/PhysRevE.80.011909
-
[15]
D. B. Saakian, T. Yakushkina, and E. V. Koonin,Allele fixation probability in a Moran model with fluctuating fit- ness landscapes,Phys. Rev. E99, 022407 (2019)
2019
-
[16]
Antal and I
T. Antal and I. Scheuring,Fixation of strategies for an evolutionary game in finite populations, Bull. Math. Biol. 68, 1923–1944 (2006)
1923
-
[17]
2017;544(7649):227– 230.doi:10.1038/nature21723
Allen B, Lippner G, Chen YT, Fotouhi B, Momeni N, Yau ST, Nowak MA.Evolutionary dynamics on any population structure.Nature. 2017;544(7649):227– 230.doi:10.1038/nature21723
-
[18]
Hindersin L, Traulsen A.Most undirected ran- dom graphs are amplifiers of selection for birth– death dynamics. PLoS Computational Biology. 2015;11(11):e1004437.doi:10.1371/journal.pcbi.1004437
-
[19]
Lieberman, C
E. Lieberman, C. Hauert, and M. A. Nowak,Evolution- ary dynamics on graphs, Nature433, 312–316 (2005)
2005
-
[20]
Ohtsuki, C
H. Ohtsuki, C. Hauert, E. Lieberman, and M. A. Nowak, A simple rule for the evolution of cooperation on graphs and social networks, Nature441, 502–505 (2006)
2006
-
[21]
1963;18:553– 556.doi:10.1038/hdy.1963.66
Ewens WJ.The probability of survival of a new mu- tant in a fluctuating environment.Heredity. 1963;18:553– 556.doi:10.1038/hdy.1963.66
-
[22]
Karlin S, Taylor HM.A First Course in Stochastic Pro- cesses.New York: Academic Press; 1975
1975
-
[23]
Ferreira EM, Neves AGM.Fixation probabilities for the Moran process with three or more strategies: general and coupling results. J. Math. Biol. 81, 277–314 (2020). https://doi.org/10.1007/s00285-020-01510-0
-
[24]
2009;75(4):266– 277.doi:10.1016/j.tpb.2009.01.009
Lessard S, Lahaie P.Fixation probability with multi- ple alleles and projected average allelic effect on se- lection.Theoretical Population Biology. 2009;75(4):266– 277.doi:10.1016/j.tpb.2009.01.009
-
[25]
Traulsen A, Claussen JC, Hauert C.Coevolutionary dynamics: From finite to infinite popu- lations. Phys Rev Lett. 2005;95:238701. doi:10.1103/PhysRevLett.95.238701
-
[26]
2006;74(1):011901.doi:10.1103/PhysRevE.74.011901
Traulsen A, Claussen JC, Hauert C.Coevolutionary dy- namics in large, but finite populations.Physical Review E. 2006;74(1):011901.doi:10.1103/PhysRevE.74.011901
-
[27]
Gardiner CW.Stochastic Methods: A Handbook for the Natural and Social Sciences.4th ed.Berlin: Springer; 2009
2009
-
[28]
2009;459:253–256.doi:10.1038/nature07921
Gore J, Youk H, van Oudenaarden A.Snowdrift game dynamics and facultative cheating in yeast.Nature. 2009;459:253–256.doi:10.1038/nature07921
-
[29]
2012;109(21):8259– 8263.doi:10.1073/pnas.1118131109
Darch SE, West SA, Winzer K, Diggle SP.Density- dependent fitness benefits in quorum-sensing bac- terial populations.Proceedings of the National Academy of Sciences USA. 2012;109(21):8259– 8263.doi:10.1073/pnas.1118131109
-
[30]
2010;331:175–178.doi:10.1126/science.1195942
Handegard NO, Boswell KM, Ioannou CC, Leblanc SP, Tjøstheim D, Couzin ID.The dynamics of coordinated group hunting and collective evasion in wild fish.Science. 2010;331:175–178.doi:10.1126/science.1195942
-
[31]
1998;102– 103:127–144.doi:10.1023/A:1017067816551
Gerrish PJ, Lenski RE.The fate of competing beneficial mutations in an asexual population.Genetica. 1998;102– 103:127–144.doi:10.1023/A:1017067816551
-
[32]
2007;176(3):1759– 1798.doi:10.1534/genetics.106.067678
Desai MM, Fisher DS.Beneficial mutation– selection balance and the effect of linkage on positive selection.Genetics. 2007;176(3):1759– 1798.doi:10.1534/genetics.106.067678
-
[33]
Toral R, Colet P.Stochastic Numerical Methods: An In- troduction for Students and Scientists.Weinheim: Wiley; 2014
2014
-
[34]
2017;548:210– 213.doi:10.1038/nature23273
Grilli J, Barab´ as G, Michalska-Smith MJ, Allesina S.Higher-order interactions stabilize dynamics in competitive network models.Nature. 2017;548:210– 213.doi:10.1038/nature23273
-
[35]
2010;81(2):021116.doi:10.1103/PhysRevE.81.021116
Assaf M, Meerson B.Extinction of metastable stochastic populations.Physical Review E. 2010;81(2):021116.doi:10.1103/PhysRevE.81.021116
-
[36]
2002;99(2):786– 790.doi:10.1073/pnas.012399899
Czaran TL, Hoekstra RF, Pagie L.Chemical warfare between microbes promotes biodiversity.Proceedings of the National Academy of Sciences. 2002;99(2):786– 790.doi:10.1073/pnas.012399899
-
[37]
Hofbauer J, Sigmund K.Evolutionary Games and Popu- lation Dynamics.Cambridge University Press. 1998
1998
-
[38]
2007;76:050901.doi:10.1103/PhysRevE.76.050901
Traulsen A, Claussen JC, Hauert C.Stochastic differ- ential equations for evolutionary dynamics with de- mographic noise and mutations.Physical Review E. 2007;76:050901.doi:10.1103/PhysRevE.76.050901
-
[39]
2014;68:109– 143.doi:10.1007/s00285-012-0604-8
Allen B, Tarnita CE.Measures of success in a class of evolutionary models with fixed population size and structure.Journal of Mathematical Biology. 2014;68:109– 143.doi:10.1007/s00285-012-0604-8. Appendix A: Fokker-Planck equation in a multi-allele Moran process We consider a Moran process withMalleles in a population of sizeN. The state of the system is ...
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.