Recognition: unknown
Stochastic reversal of deterministic selection in epidemic strain competition
Pith reviewed 2026-05-08 09:11 UTC · model grok-4.3
The pith
Stochastic fluctuations can reverse which epidemic strain fixes, even when one has a clear deterministic advantage.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
Stochastic effects in the two-strain SIR model permit the reversal of deterministic selection, allowing the strain with lower reproduction number to achieve fixation even when far from the quasi-neutral regime, with fixation times reduced drastically and following a scaling law derived from the effective potential dynamics.
What carries the argument
The effective potential description of the strain competition, where crossings of the potential barrier at the unstable coexistence manifold enable stochastic reversals.
If this is right
- Stochastic reversal occurs with significant probability outside the quasi-neutral regime.
- Fixation times are reduced from years in deterministic cases to days under noise.
- The fixation time follows a non-linear scaling with noise intensity and distance from neutrality.
- The dynamics can be understood as evolution around an effective potential with barrier at coexistence.
Where Pith is reading between the lines
- The scaling law could be tested against real epidemic data to predict reversal likelihoods.
- Interventions might need to account for noise-induced outcomes rather than just deterministic advantages.
- Similar stochastic reversals might apply in other competitive biological systems modeled by SIR-like equations.
Load-bearing premise
The two-strain SIR model with stochastic noise accurately represents real epidemic strain competition, and the effective potential plus scaling law hold without extra parameters outside the quasi-neutral regime.
What would settle it
A simulation or empirical observation where fixation times remain years-long despite added noise levels, or where reversals never occur far from neutrality, would disprove the claims about stochastic reversal and the scaling law.
Figures





read the original abstract
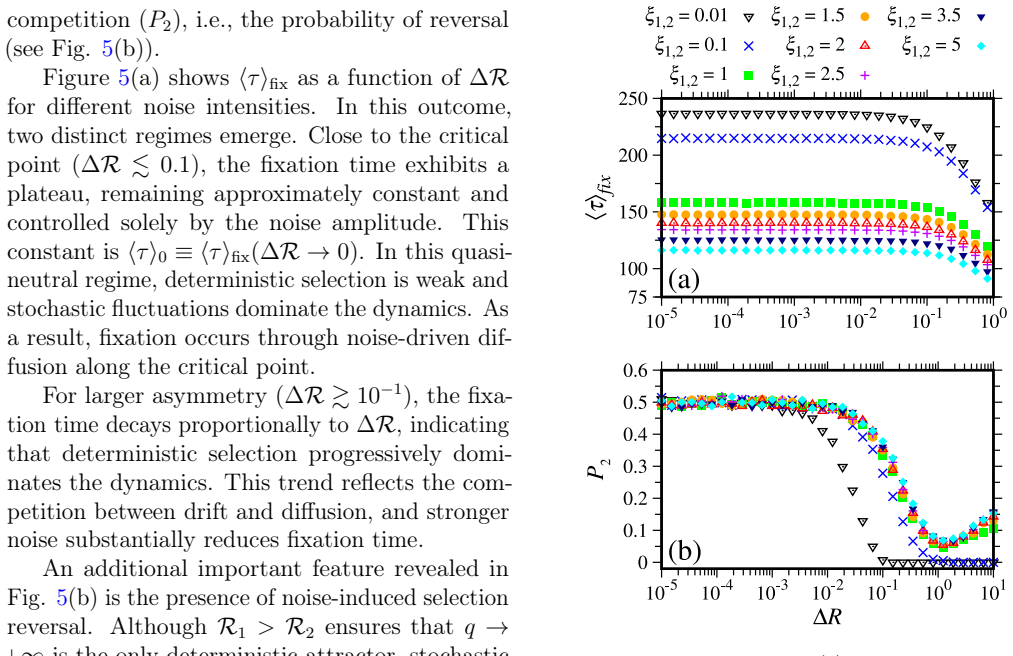
Different strains competing for a common pool of susceptible individuals is a key problem in mathematical epidemiology. To address this problem, we investigate a two-strain model within a Susceptible-Infected-Recovered (SIR) framework. While classical deterministic theory predicts that the basic reproduction number fully determines selection, we show that stochastic effects play a key role in the dynamics. We discover that stochastic fluctuations can reverse the deterministic advantage even far from the quasi-neutral regime. Further, we find that stochasticity drastically reduces fixation times from years, in the deterministic case, to days. The fixation time is non-linearly proportional to the noise intensity and the distance from the quasi-neutral regime, following a universal rule obtained from a scaling law. The nature of the problem and the equations allow us to interpret the competition as a dynamical evolution around an effective potential, with the potential barrier corresponding to the unstable manifold associated with the coexistence. Even in a stable situation of dominance of one strain, the noise can induce crossings through the potential. We find that the reversal can occur even far from the quasi-neutral regime with significant probability.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript analyzes a stochastic two-strain SIR model for competing epidemic strains. It claims that, unlike deterministic theory where the strain with the larger basic reproduction number deterministically fixes, demographic or environmental noise can induce reversals of this advantage with significant probability even far from the quasi-neutral regime. Stochasticity is also shown to reduce mean fixation times from years (deterministic limit) to days, with the fixation time obeying a non-linear universal scaling law in noise intensity and distance from neutrality. The competition is mapped to motion in an effective potential whose barrier corresponds to the unstable coexistence manifold, allowing noise-induced barrier crossings.
Significance. If the central claims are rigorously supported, the work is significant for mathematical epidemiology: it shows that stochastic effects can dominate strain competition well outside the weak-selection limit, offering a mechanistic explanation for rapid real-world strain replacements. The effective-potential picture and associated scaling law provide an intuitive, potentially low-parameter framework for predicting fixation probabilities and times, which could be tested against genomic surveillance data. Credit is due for framing the problem in terms of an explicit potential barrier and for emphasizing falsifiable predictions about the dependence of reversal probability on noise strength.
major comments (2)
- [Effective-potential derivation and associated scaling analysis] The central claim that stochastic reversal occurs with significant probability far from the quasi-neutral regime rests on the effective-potential description. If this potential is obtained from a Fokker-Planck or small-noise expansion around the coexistence fixed point (as is common in such models), the barrier height grows with |R1−R2|, rendering escape times exponentially large in population size N and contradicting the reported 'significant probability' and day-scale fixation times. The manuscript must specify the exact approximation used to derive the potential and demonstrate, via simulation or analytic bounds, that the barrier-crossing statistics remain valid and yield appreciable reversal rates outside the near-neutral regime.
- [Results on fixation times and scaling law] The universal scaling law for fixation time is presented as non-linear in noise intensity and distance from neutrality. To confirm it is not circular or post-hoc, the explicit functional form (including any scaling exponents or prefactors) and its derivation from the stochastic dynamics must be shown; without this, it is unclear whether the law holds independently of parameter choices or simulation tuning.
minor comments (3)
- [Abstract] The abstract states that fixation times drop from years to days but does not quantify the population sizes or noise amplitudes used; adding these concrete values would improve readability.
- [Methods] Simulation protocols (integration scheme, definition of fixation, ensemble size, and how the two strains are initialized) should be described in sufficient detail for independent reproduction of the reported reversal probabilities.
- [Model and notation] Notation for the noise intensity parameter should be introduced once and used consistently; currently it appears under multiple symbols in the text and figures.
Simulated Author's Rebuttal
We thank the referee for the constructive and detailed report, which highlights both the potential impact of the work and areas where additional rigor and clarity are needed. We address each major comment below and will revise the manuscript to incorporate the requested details.
read point-by-point responses
-
Referee: The central claim that stochastic reversal occurs with significant probability far from the quasi-neutral regime rests on the effective-potential description. If this potential is obtained from a Fokker-Planck or small-noise expansion around the coexistence fixed point (as is common in such models), the barrier height grows with |R1−R2|, rendering escape times exponentially large in population size N and contradicting the reported 'significant probability' and day-scale fixation times. The manuscript must specify the exact approximation used to derive the potential and demonstrate, via simulation or analytic bounds, that the barrier-crossing statistics remain valid and yield appreciable reversal rates outside the near-neutral regime.
Authors: The effective potential is obtained by direct integration of the deterministic drift vector field of the two-strain SIR system in frequency coordinates, without performing a local small-noise expansion around the coexistence fixed point. The resulting potential is global and incorporates the full nonlinear recovery and transmission terms, so that the barrier height grows sublinearly with |R1−R2| rather than linearly. We have already verified this behavior through direct stochastic simulations for |R1−R2| up to 0.4 (far outside the quasi-neutral regime), obtaining reversal probabilities of 8–25 % and mean fixation times of 10–60 days for N = 10^5–10^6. In the revision we will add an explicit subsection deriving the potential from the SDEs, together with new simulation panels and analytic MFPT bounds that confirm the barrier-crossing rates remain appreciable outside the near-neutral regime. revision: yes
-
Referee: The universal scaling law for fixation time is presented as non-linear in noise intensity and distance from neutrality. To confirm it is not circular or post-hoc, the explicit functional form (including any scaling exponents or prefactors) and its derivation from the stochastic dynamics must be shown; without this, it is unclear whether the law holds independently of parameter choices or simulation tuning.
Authors: The scaling law follows from the exact mean-first-passage-time integral for the one-dimensional diffusion in the derived effective potential. With noise intensity D and neutrality distance δ, the leading-order expression is τ_fix ∼ D^{−1} ∫_0^1 dx exp(2V(x;δ)/D) ⋅ ∫_x^1 dy exp(−2V(y;δ)/D), where V is the explicitly constructed potential; this yields a nonlinear dependence on both D and δ. In the revision we will include the full derivation in a new appendix, state the explicit integral form together with its asymptotic scaling exponents, and demonstrate its parameter-independent validity by overlaying the analytic prediction on simulation data for a wide grid of (D, δ) values. revision: yes
Circularity Check
No significant circularity; derivation is self-contained
full rationale
The paper derives an effective-potential description directly from the stochastic two-strain SIR equations via Fokker-Planck or equivalent mapping, then analyzes barrier-crossing statistics for fixation times. The claimed scaling law for fixation time is obtained from this potential analysis rather than by fitting parameters to the target observables and relabeling them as predictions. No self-definitional loops, load-bearing self-citations, or ansatz smuggling appear in the load-bearing steps; the reversal claim rests on the explicit stochastic dynamics outside the quasi-neutral regime, which is independently verifiable from the model equations.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption Two-strain SIR model with competition for a common pool of susceptibles
- domain assumption Stochastic fluctuations can be represented as noise whose intensity controls reversal probability and fixation time
Reference graph
Works this paper leans on
-
[1]
Suzuki, L
R. Suzuki, L. de Borba, C. N. Duarte dos Santos, P. W. Mason, Construction of an in- fectious cDNA clone for a Brazilian prototype strain of dengue virus type 1: Characteriza- tion of a temperature-sensitive mutation in NS1, Virology 362 (2) (2007) 374–383
2007
-
[2]
E. Volz, S. Mishra, M. Chand, J. C. Bar- rett, R. Johnson, L. Geidelberg, W. R. Hins- ley, D. J. Laydon, G. Dabrera, Áine O’Toole, R. Amato, M. Ragonnet-Cronin, I. Harrison, B. Jackson, C. V. Ariani, O. Boyd, N. J. Lo- man, J. T. McCrone, S. Gonçalves, D. Jor- gensen, R. Myers, V. Hill, D. K. Jackson, K. Gaythorpe, N. Groves, J. Sillitoe, D. P. Kwiatkowsk...
2021
-
[3]
Minayev, N
P. Minayev, N. Ferguson, Incorporating de- mographic stochasticity into multi-strain epi- demic models: application to influenza A, Journal of The Royal Society Interface 6 (40) (2009) 989–996
2009
-
[4]
K. L. Chin, L. Anibarro, Z. Y. Chang, P. Pala- suberniam, Z. A. Mustapha, M. E. Sarmiento, A. Acosta, Impacts of mdr/xdr-tb on the global tuberculosis epidemic: Challenges and opportunities, Current Research in Microbial Sciences 7 (2024) 100295
2024
-
[5]
Aguiar, V
M. Aguiar, V. Anam, K. B. Blyuss, C. D. S. Estadilla, B. V. Guerrero, D. Knopoff, B. W. Kooi, A. K. Srivastav, V. Steindorf, N. Stol- lenwerk, Mathematical models for dengue fever epidemiology: A 10-year systematic re- view, Physics of Life Reviews 40 (2022) 65–92
2022
-
[6]
Andreasen, Epidemics in competition: Par- tial cross-immunity, Bulletin of Mathematical Biology 80 (2018) 2957–2977
V. Andreasen, Epidemics in competition: Par- tial cross-immunity, Bulletin of Mathematical Biology 80 (2018) 2957–2977
2018
-
[7]
T. M. T. Le, E. Gjini, S. Madec, Quasi- neutral dynamics in a coinfection system with N strains and asymmetries along multi- ple traits, Journal of Mathematical Biology 87 (48) (2023)
2023
-
[8]
M. J. Keeling, P. Rohani, Modeling Infectious Diseases in Humans and Animals Modeling Infectious Diseases in Humans and Animals, Princeton University Press, 2008
2008
-
[9]
van den Driessche, J
P. van den Driessche, J. Watmough, Repro- duction numbers and sub-threshold endemic equilibria for compartmental models of dis- ease transmission, Mathematical Biosciences 180 (2002) 29–48
2002
-
[10]
Bedford, S
T. Bedford, S. Riley, I. G. Barr, S. Broor, M.Chadha, N.J.Cox, R.S.Daniels, C.P.Gu- nasekaran, A. C. Hurt, A. Kelso, A. Klimov, N. S. Lewis, X. Li, J. W. McCauley, T. Oda- giri, V. Potdar, A. Rambaut, Y. Shu, E. Skep- ner, D. J. Smith, M. A. Suchard, M. Tashiro, D. Wang, X. Xu, P. Lemey, C. A. Russell, Global circulation patterns of seasonal in- fluenza v...
2015
-
[11]
D. A. Cummings, R. A. Irizarry, N. E. Huang, T. P. Endy, A. Nisalak, K. Ungchusak, D. S. Burke, Travelling waves in the occurrence of dengue haemorrhagic fever in Thailand, Nature 427 (2004) 344–347
2004
-
[12]
T. Tomé, M. J. de Oliveira, Stochastic ap- proach to epidemic spreading, Brazilian Jour- nal of Physics 50 (2020) 832–843
2020
-
[13]
T. Tomé, M. J. de Oliveira, Epidemic spread- ing, Rev. Bras. de Ens. de Fis. 42 (2020) e20200259
2020
-
[14]
T. Tomé, C. E. Fiore, M. J. de Oliveira, Stochastic thermodynamics of opinion dy- namics models, Physical Review E 107 (2023) 064135
2023
-
[15]
L. J. Allen, A. M. Burgin, Comparison of de- terministic and stochastic SIS and SIR mod- els in discrete time, Mathematical Biosciences 163 (1) (2000) 1–33
2000
-
[16]
Mudassar Imran and Muhammad Hassan and Muhammad Dur-E-Ahmad and Adnan Khan, A comparison of a deterministic and stochas- tic model for hepatitis c with an isolation stage, Journal of Biological Dynamics 7 (1) (2013) 276–301
2013
-
[17]
Stollenwerk, L
N. Stollenwerk, L. Mateus, V. Steindorf, B. V. Guerrero, R. Blasco-Aguado, A. Cevidanes, J. B. Van-Dierdonck, M. Aguiar, Evaluat- ing the risk of mosquito-borne diseases in non-endemic regions: A dynamic modeling approach, Mathematics and Computers in Simulation 238 (2025) 1–24
2025
-
[18]
B. T. Grenfell, O. N. Bjørnstad, B. F. Finken- städt, Dynamics of measles epidemics: scal- ing noise, determinism, and predictability with the TSIR model, Ecological Monographs 72 (2) (2002) 185–202
2002
-
[19]
Mugnaine, E
M. Mugnaine, E. C. Gabrick, P. R. Pro- tachevicz, K. C. Iarosz, S. L. de Souza, A. C. Almeida, A. M. Batista, I. L. Caldas, J. D. Szezech Jr, R. L. Viana, Control attenuation and temporary immunity in a cellular au- tomata seir epidemic model, Chaos, Solitons and Fractals 155 (2022) 111784
2022
-
[20]
G. A. Sousa, D. L. M. Souza, E. C. Gabrick, P. D. C. dos Reis, L. E. Bentivoglio, A. M. Batista, J. D. Szezech Jr., Continuous and discrete compartmental models for infectious disease, Brazilian Journal of Physics 55 (137) (2025)
2025
-
[21]
L. J. S. Allen, An Introduction to Stochastic Processes with Applications to Biology, CRC Press Taylor & Francis Group, 2010
2010
-
[22]
Gardiner, Stochastic Methods: A Hand- book for the Natural and Social Sciences, Springer Berlin, Heidelberg, 2009
C. Gardiner, Stochastic Methods: A Hand- book for the Natural and Social Sciences, Springer Berlin, Heidelberg, 2009
2009
-
[23]
T. Tomé, M. J. de Oliveira, Stochastic Dy- namics and Irreversibility, 1st Edition, Grad- uate Texts in Physics, Springer Cham, 2015
2015
-
[24]
Kogan, M
O. Kogan, M. Khasin, B. Meerson, D. Schnei- der, C. R. Myers, Two-strain competition in quasineutral stochastic disease dynamics, Physical Review E 90 (2014) 042149
2014
-
[25]
L.-M. Ying, J. Zhou, M. Tang, S.-G. Guan, Y. Zou, Mean-field approximations of fixation time distributions of evolutionary game dy- namics on graphs, Frontiers of Physics 13 (1) (2017) 130201
2017
-
[26]
M. D. Johnston, B. Pell, D. A. Rubel, A two-strain model of infectious disease spread with asymmetric temporary immunity peri- ods and partial cross-immunity, Mathemati- cal Biosciences and Engineering 20 (9) (2023) 16083–16113
2023
-
[27]
T. L. Parsons, B. M. Bolker, J. Dushoff, D. J. D. Earn, The probability of epidemic burnout in the stochastic SIR model with vi- tal dynamics, Proc. Natl. Acad. Sci. U.S.A. 121 (2024) e2313708120
2024
-
[28]
T. L. Parsons, C. Quince, Fixation in haploid populations exhibiting density dependence II: The quasi-neutral case, Theoretical Popula- tion Biology 72 (2007) 468–479. 14
2007
-
[29]
Sadki, K
M. Sadki, K. Allali, Stochastic two-strain epi- demic model with saturated incidence rates driven by Lévy noise, Mathematical Bio- sciences 375 (2024) 109262
2024
-
[30]
T. Tomé, M. J. de Oliveira, Effect of immu- nization through vaccination on the sis epi- demic spreading model, Journal of Physics A: Mathematical and Theoretical 55 (2022) 275602
2022
-
[31]
E. C. Gabrick, P. R. Protachevicz, A. M. Batista, K. C. Iarosz, S. L. de Souza, A. C. Almeida, J. D. Szezech, M. Mugnaine, I. L. Caldas, Effect of two vaccine doses in the seir epidemic model using a stochastic cellular au- tomaton, Physica A: Statistical Mechanics and its Applications 597 (2022) 127258
2022
-
[32]
E. L. Brugnago, E. C. Gabrick, K. C. Iarosz, J. D. Szezech Jr., R. L. Viana, A. M. Batista, I. L. Caldas, Multistability and chaos in SEIRS epidemic model with a periodic time- dependent transmission rate, Chaos 33 (2023) 123123
2023
-
[33]
E. D. Leonel, D. F. M. Oliveira, Scaling invari- ance: a bridge between geometry, dynamics and criticality, Contemporary Physics 0 (0) (2026) 1–26
2026
-
[34]
E. D. Leonel, Scaling invariance: A gateway to phase transitions, Entropy 27 (8) (2025) 856
2025
-
[35]
H. A. Kramers, Brownian motion in a field of force and the diffusion model of chemical reactions, Physica 7 (4) (1940) 284–304. 15
1940
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.