Recognition: unknown
Triple Configuration of Brain Networks Based on Recurrent Neural Networks: The Synergistic Effects of Exogenous Stimuli, Task Demands, and Spontaneous Activity
Pith reviewed 2026-05-08 04:53 UTC · model grok-4.3
The pith
Recurrent neural networks can separate brain network activity into three distinct configurations driven by external stimuli, task demands, and spontaneous activity, with the parietal network as the central hub.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
The authors formalize a triple configuration framework for brain networks using an RNN trained on source-localized resting-state EEG. This framework separates latent factors of brain dynamics into those driven by exogenous stimuli, information processing tasks, and spontaneous activities. The parietal network is identified as the critical hub supporting these patterns, and anterior and posterior parietal regions show distinct functional specializations under different stimulus modalities.
What carries the argument
Recurrent neural network with neural dynamic constraints applied to source-localized EEG data, which learns to isolate and attribute distinct network configuration patterns to exogenous stimuli, task demands, and spontaneous activity.
If this is right
- The parietal network coordinates higher-order intelligence by supporting multiple simultaneous configuration patterns.
- Anterior and posterior parietal regions maintain distinct specializations that depend on the modality of incoming stimuli.
- Separating these three latent factors from neural data explains the sources of cognitive flexibility without requiring direct factor manipulation.
- Computational models constrained by neural dynamics can decode interactions among external, task, and internal influences in high-dimensional EEG.
Where Pith is reading between the lines
- The framework might be tested on task-based or stimulus-driven EEG to check whether the same triple separation emerges under direct manipulation.
- If parietal hub activity is disrupted, the model predicts measurable mixing of the three configuration patterns and corresponding drops in cognitive flexibility.
- Similar RNN approaches could be applied to other imaging modalities to test whether the triple configuration generalizes beyond EEG.
Load-bearing premise
The RNN model, trained only on resting-state EEG, can reliably isolate and attribute distinct configuration patterns to exogenous stimuli, task demands, and spontaneous activity without direct experimental manipulation of those factors.
What would settle it
Apply the trained model to new EEG recordings in which one factor is experimentally controlled, such as presenting specific stimuli or tasks, and test whether the attributed configuration patterns cleanly separate or instead become mixed.
Figures


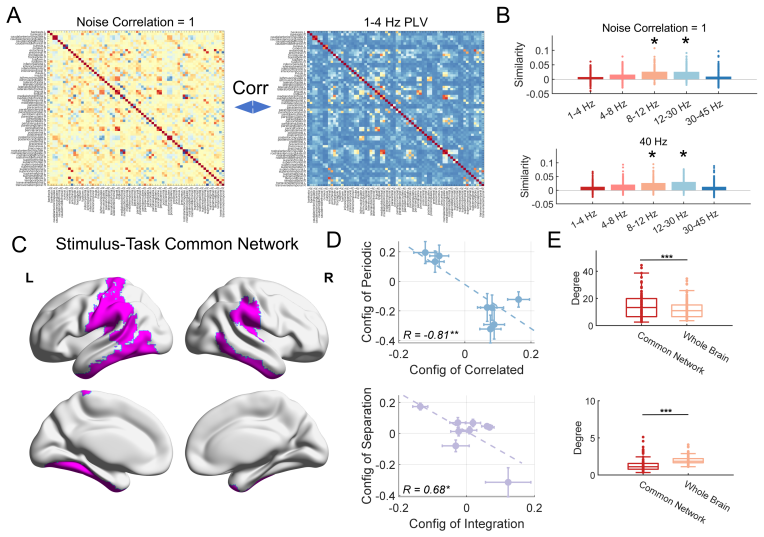


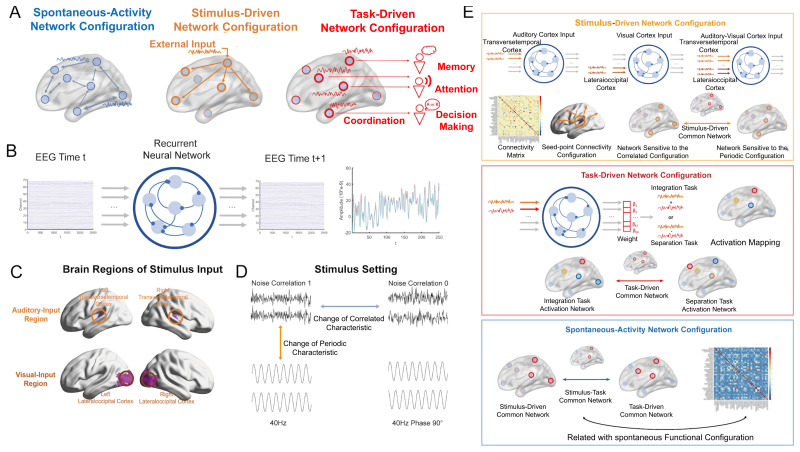


read the original abstract
The foundation of cognitive flexibility and higher-order intelligence lies in the functional structure and activity of brain networks, which can be dynamically configured by both external environments and internal states. However, decoding these dynamics from high-dimensional neural data remains a challenge. In this study, we propose a computational framework using Recurrent Neural Networks (RNNs) with neural dynamic constraints to model source-localized resting-state EEG data from $114$ participants. We aim to clarify the "triple brain network configurations" driven by exogenous and endogenous factors, including external stimuli, information processing tasks, and spontaneous activities. Our model identifies the parietal network as a critical hub supporting these multiple configuration patterns. Furthermore, we reveal that the anterior and posterior parietal regions exhibit distinct functional specializations under different stimulus modalities. By formalizing a triple configuration framework, this work separates latent factors of brain dynamics and underscores the computational significance of parietal regions in orchestrating higher-order intelligence.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript proposes a Recurrent Neural Network (RNN) framework with neural dynamic constraints, trained on source-localized resting-state EEG from 114 participants, to model and separate 'triple brain network configurations' arising from exogenous stimuli, task demands, and spontaneous activity. It claims this identifies the parietal network as a critical hub with distinct anterior-posterior specializations, thereby separating latent factors of brain dynamics and underscoring parietal contributions to higher-order intelligence.
Significance. If the separation of latent factors from spontaneous-activity data alone can be shown to be robust and non-circular, the work would provide a novel computational lens on how brain networks dynamically reconfigure for cognitive flexibility, with potential implications for understanding parietal orchestration of intelligence.
major comments (2)
- [Abstract] Abstract: The central claim that the model separates latent factors of exogenous stimuli, task demands, and spontaneous activity is load-bearing, yet the abstract supplies no equations, derivation steps, quantitative metrics, error bars, statistical tests, or baseline comparisons to support the parietal-hub identification or anterior-posterior specializations.
- [Methods] Methods (training description): The RNN is fitted exclusively to resting-state EEG, which contains only spontaneous activity. Attribution of distinct triple-configuration patterns to the two absent factors therefore rests on unverified assumptions about counterfactual modulation; this assumption directly supports the parietal-hub conclusion for higher-order intelligence and requires explicit validation or additional controls.
minor comments (2)
- [Abstract] Abstract: The phrase 'triple configuration framework' is introduced without a concise definition or pointer to the relevant equations or prior literature.
- [Discussion] The manuscript would benefit from a dedicated limitations paragraph addressing the absence of direct experimental manipulation of exogenous and task factors.
Simulated Author's Rebuttal
Thank you for the detailed review. We address the major comments point-by-point and outline revisions to enhance the manuscript's clarity and rigor.
read point-by-point responses
-
Referee: [Abstract] Abstract: The central claim that the model separates latent factors of exogenous stimuli, task demands, and spontaneous activity is load-bearing, yet the abstract supplies no equations, derivation steps, quantitative metrics, error bars, statistical tests, or baseline comparisons to support the parietal-hub identification or anterior-posterior specializations.
Authors: The abstract provides a concise summary of the study's contributions. Detailed equations, derivations, metrics, and statistical tests are presented in the Methods and Results sections of the manuscript. To address this concern, we will revise the abstract to incorporate key quantitative metrics, such as the parietal network's hub centrality score and associated p-values, while respecting length limits. This will better support the claims without equations, which are typically reserved for the main text. revision: yes
-
Referee: [Methods] Methods (training description): The RNN is fitted exclusively to resting-state EEG, which contains only spontaneous activity. Attribution of distinct triple-configuration patterns to the two absent factors therefore rests on unverified assumptions about counterfactual modulation; this assumption directly supports the parietal-hub conclusion for higher-order intelligence and requires explicit validation or additional controls.
Authors: We acknowledge that training is performed on resting-state data. The triple configurations are separated by embedding dynamic constraints that model how exogenous stimuli and task demands would alter the spontaneous dynamics, based on established neurophysiological principles. To validate this approach and demonstrate it is non-circular, we will add simulations where ground-truth modulations are applied to synthetic data, and show the model correctly attributes the configurations. We will also include baseline comparisons without constraints to confirm the separation relies on the proposed framework. revision: yes
Circularity Check
No significant circularity detected
full rationale
The abstract and available text describe a proposed RNN framework with neural dynamic constraints trained exclusively on source-localized resting-state EEG to model and separate triple brain network configurations attributed to exogenous stimuli, task demands, and spontaneous activity. No equations, derivation steps, self-citations, or explicit modeling choices are provided that reduce any claimed prediction or separation to the fitted inputs by construction. The parietal hub identification is presented as an output of the model rather than a tautological redefinition of the training data. Without load-bearing steps that can be quoted and shown to equate outputs to inputs, the derivation chain is self-contained as a computational modeling proposal.
Axiom & Free-Parameter Ledger
Reference graph
Works this paper leans on
-
[1]
Selective entrainment of theta oscillations in the dorsal stream causally enhances auditory working memory performance
Philippe Albouy, Avi Weiss, Sylvain Baillet, and Robert J Zatorre. Selective entrainment of theta oscillations in the dorsal stream causally enhances auditory working memory performance. Neuron, 94(1):193–206, 2017
2017
-
[2]
Supramodality of neural entrainment: Rhythmic visual stimulation causally enhances auditory working memory performance.Science advances, 8(15):eabj9782, 2022
Philippe Albouy, Mélanie Rufiange, Isabelle Peretz, and Robert J Zatorre. Supramodality of neural entrainment: Rhythmic visual stimulation causally enhances auditory working memory performance.Science advances, 8(15):eabj9782, 2022
2022
-
[3]
Effects of feature-selective attention on auditory pattern and location processing.Neuroimage, 42(1):69–79, 2008
Christian F Altmann, Marion Petton, Lutz Jäncke, and Jochen Kaiser. Effects of feature-selective attention on auditory pattern and location processing.Neuroimage, 42(1):69–79, 2008. 9
2008
-
[4]
Communication dynamics in complex brain networks.Nature reviews neuroscience, 19(1):17–33, 2018
Andrea Avena-Koenigsberger, Bratislav Misic, and Olaf Sporns. Communication dynamics in complex brain networks.Nature reviews neuroscience, 19(1):17–33, 2018
2018
-
[5]
A note on the phase locking value and its properties.Neuroimage, 74:231–244, 2013
Sergul Aydore, Dimitrios Pantazis, and Richard M Leahy. A note on the phase locking value and its properties.Neuroimage, 74:231–244, 2013
2013
-
[6]
Dynamic reconfiguration of human brain networks during learning
Danielle S Bassett, Nicholas F Wymbs, Mason A Porter, Peter J Mucha, Jean M Carlson, and Scott T Grafton. Dynamic reconfiguration of human brain networks during learning. Proceedings of the National Academy of Sciences, 108(18):7641–7646, 2011
2011
-
[7]
At the intersection of attention and mem- ory: the mechanistic role of the posterior parietal lobe in working memory.Neuropsychologia, 49(5):1306–1315, 2011
Marian E Berryhill, Jason Chein, and Ingrid R Olson. At the intersection of attention and mem- ory: the mechanistic role of the posterior parietal lobe in working memory.Neuropsychologia, 49(5):1306–1315, 2011
2011
-
[8]
Effects of selective attention on the electrophysiological representation of concurrent sounds in the human auditory cortex
Aurélie Bidet-Caulet, Leon Y Deouell, and Robert T Knight. Effects of selective attention on the electrophysiological representation of concurrent sounds in the human auditory cortex. Journal of Neuroscience, 27(34):9252–9261, 2007
2007
-
[9]
Attention, intention, and priority in the parietal lobe
James W Bisley and Michael E Goldberg. Attention, intention, and priority in the parietal lobe. Annual review of neuroscience, 33:1–21, 2010
2010
-
[10]
Complex brain networks: graph theoretical analysis of structural and functional systems.Nature reviews neuroscience, 10(3):186–198, 2009
Ed Bullmore and Olaf Sporns. Complex brain networks: graph theoretical analysis of structural and functional systems.Nature reviews neuroscience, 10(3):186–198, 2009
2009
-
[11]
Emergent complex neural dynamics.Nature physics, 6(10):744–750, 2010
Dante R Chialvo. Emergent complex neural dynamics.Nature physics, 6(10):744–750, 2010
2010
-
[12]
Eeglab: an open source toolbox for analysis of single-trial eeg dynamics including independent component analysis.Journal of neuroscience methods, 134(1):9–21, 2004
Arnaud Delorme and Scott Makeig. Eeglab: an open source toolbox for analysis of single-trial eeg dynamics including independent component analysis.Journal of neuroscience methods, 134(1):9–21, 2004
2004
-
[13]
Posterior parietal cortex dysfunction is central to working memory storage and broad cognitive deficits in schizophrenia.Journal of Neuroscience, 38(39):8378–8387, 2018
Britta Hahn, Benjamin M Robinson, Stefan T Kaiser, Alex N Harvey, Valerie M Beck, Carlyle J Leonard, James M Gold, and Steven J Luck. Posterior parietal cortex dysfunction is central to working memory storage and broad cognitive deficits in schizophrenia.Journal of Neuroscience, 38(39):8378–8387, 2018
2018
-
[14]
Graph theory approaches to functional network organization in brain disorders: A critique for a brave new small-world.Network neuroscience, 3(1):1–26, 2018
Michael N Hallquist and Frank G Hillary. Graph theory approaches to functional network organization in brain disorders: A critique for a brave new small-world.Network neuroscience, 3(1):1–26, 2018
2018
-
[15]
Scale-free brain activity: past, present, and future.Trends in cognitive sciences, 18 (9):480–487, 2014
Biyu J He. Scale-free brain activity: past, present, and future.Trends in cognitive sciences, 18 (9):480–487, 2014
2014
-
[16]
Dynamic categorization rules alter representations in human visual cortex.Nature Communications, 16 (1):3459, 2025
Margaret M Henderson, John T Serences, and Nuttida Rungratsameetaweemana. Dynamic categorization rules alter representations in human visual cortex.Nature Communications, 16 (1):3459, 2025
2025
-
[17]
Measuring the dynamic balance of integration and segregation underlying consciousness, anesthesia, and sleep in humans.Nature communications, 15(1):9164, 2024
Hyunwoo Jang, Minkyung Lee, Sel-In Kim, Hang-Kwon Kim, Seong-Wook Kim, Seung-Woo Lee, George A Mashour, and Uncheol Lee. Measuring the dynamic balance of integration and segregation underlying consciousness, anesthesia, and sleep in humans.Nature communications, 15(1):9164, 2024
2024
-
[18]
Age-related shifts in brain activity dynamics during task switching.Cerebral Cortex, 20(6):1420–1431, 2010
Koji Jimura and Todd S Braver. Age-related shifts in brain activity dynamics during task switching.Cerebral Cortex, 20(6):1420–1431, 2010
2010
-
[19]
Cortical activations during the mental rotation of different visual objects.Neuroimage, 13(1):143–152, 2001
Kirsten Jordan, Hans-Jochen Heinze, Kai Lutz, Martin Kanowski, and Lutz Jäncke. Cortical activations during the mental rotation of different visual objects.Neuroimage, 13(1):143–152, 2001
2001
-
[20]
Functional brain network reconfiguration during learning in a dynamic environment.Nature communications, 11(1):1682, 2020
Chang-Hao Kao, Po-Hsuan Chen, Fong-Hao Su, Yi-Tzu Wu, Philip Tseng, and Fa-Hsuan Lin. Functional brain network reconfiguration during learning in a dynamic environment.Nature communications, 11(1):1682, 2020
2020
-
[21]
Dynamic spatial coding in parietal cortex mediates tactile-motor transformation
Janina Klautke, Jean-Charles Pesnot-Lerousseau, Victor de Lafuente, Thomas Brochier, and Alexa Riehle. Dynamic spatial coding in parietal cortex mediates tactile-motor transformation. Nature Communications, 14(1):4532, 2023. 10
2023
-
[22]
Prefrontal transthalamic uncertainty processing drives flexible switching.Nature, 637(8000):127–136, 2025
Norman H Lam, Khiem Nguyen, Khoi D Le, Jina Chung, Charlie Nam, Soyun Choi, and Daesoo Kim. Prefrontal transthalamic uncertainty processing drives flexible switching.Nature, 637(8000):127–136, 2025
2025
-
[23]
Amygdala engages non- emotional multi-item working memory maintenance through amygdala-hippocampus interaction
Chenyang Li, Zeping Zhu, Yuhua Luo, Xiaobing Zhang, and Jin Li. Amygdala engages non- emotional multi-item working memory maintenance through amygdala-hippocampus interaction. bioRxiv, pages 2024–05, 2024
2024
-
[24]
Functional specialization and interaction in the amygdala-hippocampus circuit during working memory processing.Nature Communications, 14(1):2921, 2023
Jin Li, Zeping Zhu, Yuhua Luo, Xiaobing Zhang, Chenyang Li, Dan Zhang, and Li Yao. Functional specialization and interaction in the amygdala-hippocampus circuit during working memory processing.Nature Communications, 14(1):2921, 2023
2023
-
[25]
Transitions in information processing dynamics at the whole-brain network level are driven by alterations in neural gain.PLoS computational biology, 15(10):e1006957, 2019
Mike Li, Ben J Cruz, Joaquín Goñi, Ervin Shitiri, José Vizuete, Andri C Tziortzi, Olaf Sporns, Andreas A Ioannides, Shitij Kapur, and Meera Alle. Transitions in information processing dynamics at the whole-brain network level are driven by alterations in neural gain.PLoS computational biology, 15(10):e1006957, 2019
2019
-
[26]
Yun Li, Hui Liu, Ming Li, Weinan Li, Yajuan Yang, Tian-tian Li, Andrew E Hudson, Hong Yu, and Xiao-li Li. Comparison of nrem sleep and intravenous sedation through local information processing and whole brain network to explore the mechanism of general anesthesia.PloS one, 13(6):e0192358, 2018
2018
-
[27]
Backpropagation through time and the brain.Current opinion in neurobiology, 55:82–89, 2019
Timothy P Lillicrap and Adam Santoro. Backpropagation through time and the brain.Current opinion in neurobiology, 55:82–89, 2019
2019
-
[28]
Flexible gating between subspaces in a neural network model of internally guided task switching.Nature communications, 16(1):6497, 2025
Yue Liu and Xiao-Jing Wang. Flexible gating between subspaces in a neural network model of internally guided task switching.Nature communications, 16(1):6497, 2025
2025
-
[29]
Emergent scale-free networks.PNAS nexus, 3(3):pgad236, 2024
Christopher W Lynn, Caroline M Holmes, and Stephanie E Palmer. Emergent scale-free networks.PNAS nexus, 3(3):pgad236, 2024
2024
-
[30]
The flexible brain.Brain, 139(7): 2110–2112, 2016
Marcelo G Mattar, Richard F Betzel, and Danielle S Bassett. The flexible brain.Brain, 139(7): 2110–2112, 2016
2016
-
[31]
Functional alignment with anatomical networks is associated with cognitive flexibility.Nature human behaviour, 2(2):156–164, 2018
John D Medaglia, Fabio Pasqualetti, Roy H Hamilton, Sharon L Thompson-Schill, and Danielle S Bassett. Functional alignment with anatomical networks is associated with cognitive flexibility.Nature human behaviour, 2(2):156–164, 2018
2018
-
[32]
Computational memory capacity predicts aging and cognitive decline.Nature communi- cations, 16(1):2748, 2025
Mite Mijalkov, Ludvig Storm, Blanca Zufiria-Gerbolés, Kilian Abellaneda-Pérez, Karen Ritchie, Aida Carrié-I-Crespo, Laurence Annet, Bruno Dubois, Didier Dormont, Stéphane Epelbaum, et al. Computational memory capacity predicts aging and cognitive decline.Nature communi- cations, 16(1):2748, 2025
2025
-
[33]
Temporally and function- ally distinct large-scale brain network dynamics supporting task switching.Neuroimage, 257: 119126, 2022
Takumi Mitsuhashi, Koki Okita, Kenji Matsumoto, and Yoko Hoshi. Temporally and function- ally distinct large-scale brain network dynamics supporting task switching.Neuroimage, 257: 119126, 2022
2022
-
[34]
Pytorch: An imperative style, high-performance deep learning library.Advances in neural information processing systems, 32, 2019
Adam Paszke, Sam Gross, Francisco Massa, Adam Lerer, James Bradbury, Gregory Chanan, Trevor Killeen, Zeming Lin, Natalia Gimelshein, Luca Antiga, et al. Pytorch: An imperative style, high-performance deep learning library.Advances in neural information processing systems, 32, 2019
2019
-
[35]
Brain networks and cognitive architectures.Neuron, 88(1): 207–219, 2015
Steven E Petersen and Olaf Sporns. Brain networks and cognitive architectures.Neuron, 88(1): 207–219, 2015
2015
-
[36]
An investi- gation of the phase locking index for measuring of interdependency of cortical source signals recorded in the eeg.Biological cybernetics, 100(2):129–146, 2009
Andrei V Sazonov, Marion Müller, Thomas König, and Inga Griskova-Bulanova. An investi- gation of the phase locking index for measuring of interdependency of cortical source signals recorded in the eeg.Biological cybernetics, 100(2):129–146, 2009
2009
-
[37]
Brain network communication: concepts, models and applications.Nature reviews neuroscience, 24(9):557–574, 2023
Caio Seguin, Olaf Sporns, and Andrew Zalesky. Brain network communication: concepts, models and applications.Nature reviews neuroscience, 24(9):557–574, 2023
2023
-
[38]
Principles of dynamic network reconfiguration across diverse brain states.NeuroImage, 180:396–405, 2018
James M Shine and Russell A Poldrack. Principles of dynamic network reconfiguration across diverse brain states.NeuroImage, 180:396–405, 2018. 11
2018
-
[39]
Hub overload and failure as a final common pathway in neurological brain network disorders.Network Neuroscience, pages 1–23, 2024
Cornelis Jan Stam. Hub overload and failure as a final common pathway in neurological brain network disorders.Network Neuroscience, pages 1–23, 2024
2024
-
[40]
Structure learning and the posterior parietal cortex.Progress in neurobiology, 184:101717, 2020
Christopher Summerfield, Fabrice Luyckx, and Hannah Sheahan. Structure learning and the posterior parietal cortex.Progress in neurobiology, 184:101717, 2020
2020
-
[41]
Brainstorm: A user-friendly application for meg/eeg analysis.Computational intelligence and neuroscience, 2011, 2011
François Tadel, Sylvain Baillet, John C Mosher, Dimitrios Pantazis, and Richard M Leahy. Brainstorm: A user-friendly application for meg/eeg analysis.Computational intelligence and neuroscience, 2011, 2011
2011
-
[42]
Efficiency of a’small-world’brain network depends on consciousness level: a resting-state fmri study.Cerebral cortex, 24(6):1529–1539, 2014
Taira Uehara, Takao Yamasaki, Toshiaki Okamoto, Takahiko Koike, Shinya Kan, Satoru Miyauchi, and Shozo Tobimatsu. Efficiency of a’small-world’brain network depends on consciousness level: a resting-state fmri study.Cerebral cortex, 24(6):1529–1539, 2014
2014
-
[43]
Network hubs in the human brain.Trends in cognitive sciences, 17(12):683–696, 2013
Martijn P Van den Heuvel and Olaf Sporns. Network hubs in the human brain.Trends in cognitive sciences, 17(12):683–696, 2013
2013
-
[44]
Organizing space through saccades and fixations between primate posterior parietal cortex and hippocampus
Marie-E Vericel, Thomas Michelet, Igor Kagan, and Suliann Ben Hamed. Organizing space through saccades and fixations between primate posterior parietal cortex and hippocampus. Nature Communications, 15(1):10448, 2024
2024
-
[45]
Collective dynamics of ‘small-world’networks.nature, 393(6684):440–442, 1998
Duncan J Watts and Steven H Strogatz. Collective dynamics of ‘small-world’networks.nature, 393(6684):440–442, 1998
1998
-
[46]
The role of the posterior parietal cortex on cognition: an exploratory study.Brain Research, 1769:147452, 2021
Marlee Whybird, Melissa Johnson, and Ericka Johnson. The role of the posterior parietal cortex on cognition: an exploratory study.Brain Research, 1769:147452, 2021
2021
-
[47]
The neural signature of psychomotor disturbance in depression.Molecular psychiatry, 29(1):317–326, 2024
Florian Wüthrich, Werner Strik, Yosuke Morishima, Michael Kaess, Christoph M Michel, and Thomas J Müller. The neural signature of psychomotor disturbance in depression.Molecular psychiatry, 29(1):317–326, 2024
2024
-
[48]
Yaoda Xu. The human posterior parietal cortices orthogonalize the representation of different streams of information concurrently coded in visual working memory.PLoS biology, 22(6): e3002915, 2024
2024
-
[49]
Transformation of acoustic information to sensory decision variables in the parietal cortex.Proceedings of the National Academy of Sciences, 120(15):e2212120120, 2023
Justin D Yao, Cheng Wang, Chandramouli Chandrasekaran, Michael N Shadlen, and William T Newsome. Transformation of acoustic information to sensory decision variables in the parietal cortex.Proceedings of the National Academy of Sciences, 120(15):e2212120120, 2023
2023
-
[50]
Anatomical substrates of the alerting, orienting and executive control components of attention: focus on the posterior parietal lobe.PloS one, 7(11): e50590, 2012
Xuntao Yin, Tao Wang, and Antao Chen. Anatomical substrates of the alerting, orienting and executive control components of attention: focus on the posterior parietal lobe.PloS one, 7(11): e50590, 2012. A Detailed Methods A.1 Participants, EEG Recording, and Preprocessing A total of 114 participants (60 males and 54 females, mean age ± SE = 25.2±2.4 ) were...
2012
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.