Recognition: unknown
Equation Learning for multiscale models of infectious diseases
Pith reviewed 2026-05-07 17:10 UTC · model grok-4.3
The pith
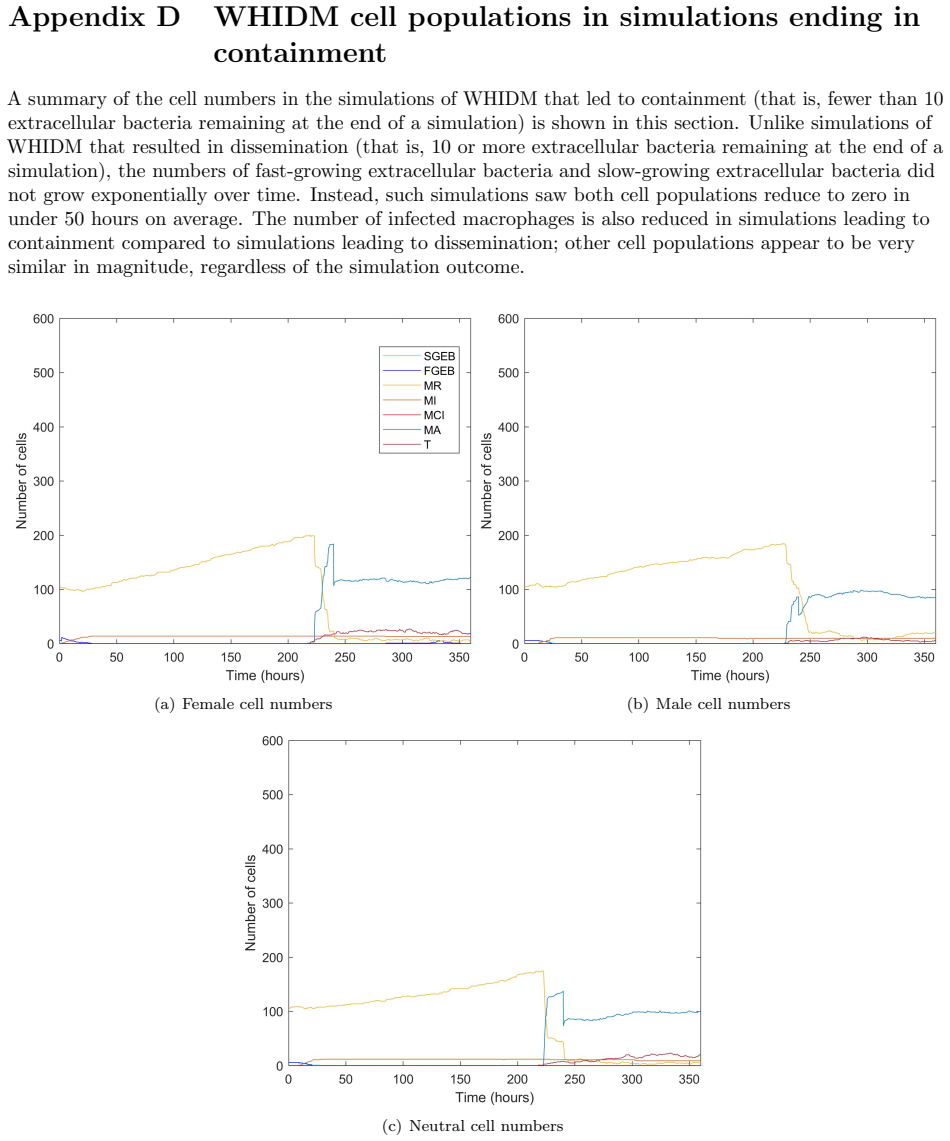
A multiscale tuberculosis model learns ODEs from agent-based simulations to link within-host immune dynamics with population-level spread and sex differences.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
We have developed a gender/sex-stratified multiscale framework for tuberculosis. We have learnt ordinary differential equations to capture the average output of an agent-based within-host model and used the resulting equations to describe the within-host scales of the multiscale framework. We evolve the population demographics at the between-host scale using ODEs and link the scales with stochastic coupling functions. We have considered counterfactual scenarios to elucidate the impact of sex and gender on the infectious disease dynamics of TB. This paper is intended to provide a proof-of-concept for the development and implementation of the presented multiscale framework.
What carries the argument
Ordinary differential equations learned from the average output of an agent-based within-host model, which are then embedded as the within-host component of the larger multiscale structure.
If this is right
- The framework can separate the contribution of within-host immune differences from population-level treatment completion rates to the observed male excess in TB cases.
- Stochastic coupling functions transmit variability between scales while keeping the overall simulation tractable.
- Counterfactual runs can identify whether altering male treatment adherence or immune parameters produces larger shifts in epidemic size.
- Replacing the agent-based within-host component with its learned ODE equivalent reduces computational cost enough to explore many demographic scenarios.
Where Pith is reading between the lines
- The same learning step could be repeated for other pathogens once an agent-based within-host model exists, yielding reusable within-host modules for new multiscale studies.
- If the learned equations remain accurate under parameter changes not seen during training, the framework could test interventions such as sex-targeted vaccines before they are trialed.
Load-bearing premise
The learned ODEs accurately represent the average dynamics of the agent-based within-host model with enough fidelity that they can be substituted into the multiscale framework without distorting the coupled population-level predictions.
What would settle it
A side-by-side run of the full agent-based within-host model inside the coupled framework versus the learned-ODE version, followed by a large mismatch in predicted sex-stratified TB incidence at the population level, would show the substitution fails.
Figures


















read the original abstract
Tuberculosis (TB) is an airborne disease caused by the pathogen Mycobacterium tuberculosis. In 2023, according to the World Health Organization, it ''probably'' replaced COVID-19 as the leading cause of death from an infectious agent globally; in the nineteenth century, one in seven of all humans deaths were as a result of tuberculosis. More than 10 million people are diagnosed with TB every year. The majority of cases in adults occur in males (62.5% of all global adult cases in 2023, compared to 37.5% in females). The main reasons for males suffering from a higher burden of global TB cases, compared to females, is likely to be a combination of within-host factors, such as differences in immune response, and population-scale factors, such as likelihood of completing treatment. To investigate the impact different scales have in determining this higher TB burden in males, we have developed a gender/sex-stratified multiscale framework. We have learnt ordinary differential equations (ODEs) to capture the average output of an agent-based within-host model, and used the resulting equations to describe the within-host scales of the multiscale framework. We evolve the population demographics at the between-host scale using ODEs, and link the scales with stochastic coupling functions. We have considered counterfactual scenarios to elucidate the impact of sex and gender on the infectious disease dynamics of TB. This paper is intended to provide a proof-of-concept for the development and implementation of the presented multiscale framework.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper develops a gender/sex-stratified multiscale framework for tuberculosis (TB) to investigate factors behind higher male burden. It learns ODEs via equation learning to approximate average trajectories from a sex-stratified agent-based within-host model, substitutes these into the within-host component of the multiscale model, evolves between-host population demographics with ODEs, and links scales via stochastic coupling functions. Counterfactual scenarios are then used to separate within-host (e.g., immune response) and population-scale (e.g., treatment completion) contributions; the work is framed as a proof-of-concept.
Significance. If the learned ODEs are shown to reproduce ABM averages with quantified fidelity and the stochastic couplings preserve key dynamics, the framework would offer a practical template for embedding detailed within-host biology into population-level infectious-disease models without prohibitive computational cost. The explicit sex/gender stratification and counterfactual design address a documented epidemiological disparity and could inform targeted interventions once validated.
major comments (2)
- [Methods (equation-learning and multiscale coupling subsections)] The central substitution step—that learned ODEs can replace the agent-based within-host model—requires demonstration that approximation error is small enough not to distort multiscale predictions. No error metrics (RMSE, R², or ensemble trajectory overlays on bacterial load or immune-cell counts), parameter regimes tested, or sensitivity checks on how residual mismatch propagates through the stochastic coupling functions are reported. This directly undermines in the counterfactual results on sex/gender effects.
- [Results (counterfactual scenarios)] The counterfactual scenarios are presented as the primary application, yet the manuscript supplies neither the specific parameter shifts used to isolate within-host versus between-host contributions nor any quantitative outcomes (incidence ratios, burden differences) or robustness tests against the within-host approximation. Without these, it is impossible to assess whether the framework successfully disentangles scales.
minor comments (2)
- [Multiscale framework description] The stochastic coupling functions are described at a high level; an explicit mathematical definition (including how noise is sampled and how state variables are mapped between scales) would improve reproducibility.
- [Figures] Figure captions and axis labels for any within-host trajectory comparisons or population-level outputs should explicitly state the number of ABM realizations averaged and the time window used for equation learning.
Simulated Author's Rebuttal
We thank the referee for their constructive and detailed comments, which highlight important aspects for strengthening our proof-of-concept multiscale framework. We address each major comment below and will revise the manuscript accordingly.
read point-by-point responses
-
Referee: [Methods (equation-learning and multiscale coupling subsections)] The central substitution step—that learned ODEs can replace the agent-based within-host model—requires demonstration that approximation error is small enough not to distort multiscale predictions. No error metrics (RMSE, R², or ensemble trajectory overlays on bacterial load or immune-cell counts), parameter regimes tested, or sensitivity checks on how residual mismatch propagates through the stochastic coupling functions are reported. This directly undermines in the counterfactual results on sex/gender effects.
Authors: We agree that explicit quantification of the learned ODE approximation error is necessary to support the substitution into the multiscale model and to lend credibility to the counterfactual findings. The current manuscript presents the overall framework conceptually as a proof-of-concept and therefore omits these metrics. In the revision we will add RMSE, R², and ensemble trajectory comparisons for bacterial load and immune-cell counts across the tested parameter regimes. We will also include sensitivity analyses showing propagation of residual mismatch through the stochastic coupling functions. These will appear in the Methods section with supporting figures. revision: yes
-
Referee: [Results (counterfactual scenarios)] The counterfactual scenarios are presented as the primary application, yet the manuscript supplies neither the specific parameter shifts used to isolate within-host versus between-host contributions nor any quantitative outcomes (incidence ratios, burden differences) or robustness tests against the within-host approximation. Without these, it is impossible to assess whether the framework successfully disentangles scales.
Authors: We accept that the counterfactual scenarios, as the main demonstration of the framework, need explicit parameter values and quantitative results to allow readers to evaluate scale disentanglement. The present version describes the scenarios at a conceptual level. In the revised manuscript we will state the precise parameter shifts used for within-host (immune-response) and between-host (treatment-completion) factors, report quantitative outcomes such as incidence ratios and sex-specific burden differences, and add robustness checks against the within-host ODE approximation. These details will be placed in the Results section together with tables and figures. revision: yes
Circularity Check
No circularity: equation learning derives ODEs from independent ABM simulations
full rationale
The derivation begins with an external agent-based within-host model whose trajectories are generated independently; ODEs are then learned to match the average output of those simulations. These learned ODEs are substituted into the multiscale framework alongside separate between-host ODEs and stochastic coupling functions. No step reduces a claimed prediction or result to a quantity defined by the same fitted parameters, nor does any load-bearing premise rest on a self-citation chain that itself lacks external verification. The construction is therefore self-contained against the ABM benchmark and does not exhibit any of the enumerated circular patterns.
Axiom & Free-Parameter Ledger
Reference graph
Works this paper leans on
-
[1]
The replication-transmission relativity theory for multiscale modelling of infectious disease systems
Garira W. The replication-transmission relativity theory for multiscale modelling of infectious disease systems. Scientific Reports 2019; 9
2019
-
[2]
Crossing the scale from within-host infection dynamics to between-host transmission fitness: a discussion of current assumptions and knowledge
Handel A and Rohani P. Crossing the scale from within-host infection dynamics to between-host transmission fitness: a discussion of current assumptions and knowledge. Philosophical Transactions of the Royal Society B: Biological Sciences 2015; 370
2015
-
[3]
Linked within-host and between-host models and data for infectious diseases: a systematic review
Childs LM, El Moustaid F, Gajewski Z, Kadelka S, Nikin-Beers R, Smith Jr JW, Walker M, and Johnson LR. Linked within-host and between-host models and data for infectious diseases: a systematic review. PeerJ 2019; 7
2019
-
[4]
A primer on multiscale modelling of infectious disease systems
Garira W. A primer on multiscale modelling of infectious disease systems. Infectious Disease Modelling 2018; 3
2018
-
[5]
The research and development process for multiscale models of infectious disease systems
Garira W. The research and development process for multiscale models of infectious disease systems. PLoS computational biology 2020; 16
2020
-
[6]
Mathematical methods for scaling from within-host to population-scale in infectious disease systems
Doran JWG, Thompson RN, Yates CA, and Bowness R. Mathematical methods for scaling from within-host to population-scale in infectious disease systems. Epidemics 2023
2023
-
[7]
Global tuberculosis report 2024
World Health Organization. Global tuberculosis report 2024. 2024. Available from: https://iris.who.int/bitstream/handle/10665/379339/9789240101531-eng.pdf?sequence=1
2024
-
[8]
Modelling challenges in context: lessons from malaria, HIV, and tuberculosis
Childs LM, Abuelezam NN, Dye C, Gupta S, Murray MB, Williams BG, and Buckee CO. Modelling challenges in context: lessons from malaria, HIV, and tuberculosis. Epidemics 2015; 10
2015
-
[9]
Pathogenesis, immunology, and diagnosis of latent Mycobacterium tuberculosis infection
Ahmad S. Pathogenesis, immunology, and diagnosis of latent Mycobacterium tuberculosis infection. Clinical and Developmental Immunology 2011; 2011
2011
-
[10]
Global Epidemiology of Tuberculosis.Seminars in Respiratory and Critical Care Medicine
Glaziou P, Floyd K, and Raviglione MC. Global Epidemiology of Tuberculosis.Seminars in Respiratory and Critical Care Medicine. Vol. 39. 3. 2018
2018
-
[11]
Modelling the effects of bacterial cell state and spatial location on tuberculosis treatment: Insights from a hybrid multiscale cellular automaton model
Bowness R, Chaplain MA, Powathil GG, and Gillespie SH. Modelling the effects of bacterial cell state and spatial location on tuberculosis treatment: Insights from a hybrid multiscale cellular automaton model. Journal of Theoretical Biology 2018; 446. Available from: https://www.sciencedirect.com/science/article/pii/S0022519318301206
2018
-
[12]
The immune response in tuberculosis
O’Garra A, Redford PS, McNab FW, Bloom CI, Wilkinson RJ, and Berry MPR. The immune response in tuberculosis. Annual review of immunology 2013; 31
2013
-
[13]
The host immune response to tuberculosis
Schluger NW and Rom WN. The host immune response to tuberculosis. American journal of respiratory and critical care medicine 1998; 157
1998
-
[14]
Phenotypic resistance in mycobacteria: is it because I am old or fat that I resist you? Journal of Antimicrobial Chemotherapy 2015; 70
Hammond RJH, Baron VO, Oravcova K, Lipworth S, and Gillespie SH. Phenotypic resistance in mycobacteria: is it because I am old or fat that I resist you? Journal of Antimicrobial Chemotherapy 2015; 70
2015
-
[15]
Defining dormancy in mycobacterial disease
Lipworth S, Hammond R, Baron V, Hu Y, Coates A, and Gillespie S. Defining dormancy in mycobacterial disease. Tuberculosis 2016; 99. Available from: https://www.sciencedirect.com/science/article/pii/S1472979216300658
2016
-
[16]
A virtual host model of Mycobacterium tuberculosis infection identifies early immune events as predictive of infection outcomes
Joslyn LR, Linderman JJ, and Kirschner DE. A virtual host model of Mycobacterium tuberculosis infection identifies early immune events as predictive of infection outcomes. Journal of theoretical biology 2022; 539
2022
-
[17]
Biological differences between the sexes and susceptibility to tuberculosis
Nhamoyebonde S and Leslie A. Biological differences between the sexes and susceptibility to tuberculosis. The Journal of infectious diseases 2014
2014
-
[18]
Genetic and hormonal mechanisms underlying sex-specific immune responses in tuberculosis
Gupta M, Srikrishna G, Klein SL, and Bishai WR. Genetic and hormonal mechanisms underlying sex-specific immune responses in tuberculosis. Trends in immunology 2022; 43
2022
-
[19]
Testosterone in pathogenesis of tuberculosis
Shrivastava P and Bagchi T. Testosterone in pathogenesis of tuberculosis. Chemical Biology Letters 2021; 8
2021
-
[20]
Predictors of Pulmonary Cavitation among Tuberculosis Patients
Balogun OO, Fawole A, Osemwinyen E, and Balogun B. Predictors of Pulmonary Cavitation among Tuberculosis Patients. J Infect Dis Epidemiol 2021; 7 31
2021
-
[21]
Cavitary tuberculosis: the gateway of disease transmission
Urbanowski ME, Ordonez AA, Ruiz-Bedoya CA, Jain SK, and Bishai WR. Cavitary tuberculosis: the gateway of disease transmission. The Lancet Infectious Diseases 2020; 20
2020
-
[22]
Risk of tuberculosis in public transport sector workers, Lima, Peru
Horna-Campos OJ, Bedoya-Lama A, Romero-Sandoval NC, and Martin-Mateo M. Risk of tuberculosis in public transport sector workers, Lima, Peru. The international journal of tuberculosis and lung disease 2010; 14
2010
-
[23]
Barriers and delays in tuberculosis diagnosis and treatment services: does gender matter? Tuberculosis research and treatment 2014; 2014
Yang WT, Gounder CR, Akande T, De Neve JW, McIntire KN, Chandrasekhar A, Lima Pereira A de, Gummadi N, Samanta S, and Gupta A. Barriers and delays in tuberculosis diagnosis and treatment services: does gender matter? Tuberculosis research and treatment 2014; 2014
2014
-
[24]
Gender differences in delays in diagnosis and treatment of tuberculosis
Karim F, Islam MA, Chowdhury AMR, Johansson E, and Diwan VK. Gender differences in delays in diagnosis and treatment of tuberculosis. Health policy and planning 2007; 22
2007
-
[25]
An assessment of factors contributing to treatment adherence and knowledge of TB transmission among patients on TB treatment
Kaona FAD, Tuba M, Siziya S, and Sikaona L. An assessment of factors contributing to treatment adherence and knowledge of TB transmission among patients on TB treatment. BMC Public health 2004; 4
2004
-
[26]
A systematic review on the role of gender in tuberculosis control
Hof S van den, Najlis CA, Bloss E, and Straetemans M. A systematic review on the role of gender in tuberculosis control. Report prepared for Tuberculosis Control Programme (TB CAP) September 2010
2010
-
[27]
A nested model for tuberculosis: Combining within-host and between-host processes in a single framework
Pereira RS, Bauch CT, Penna TJP, and Esp´ ındola AL. A nested model for tuberculosis: Combining within-host and between-host processes in a single framework. International Journal of Modern Physics C 2021; 32
2021
-
[28]
Modeling of gender differences in tuberculosis prevalence
Kisselevskaya-Babinina VY, Sannikova TE, Romanyukha AA, and Karkach AS. Modeling of gender differences in tuberculosis prevalence. Matematicheskaya Biologiya i Bioinformatika 2018; 13
2018
-
[29]
Drivers of sex differences in the South African adult tuberculosis incidence and mortality trends, 1990–2019
Kubjane M, Cornell M, Osman M, Boulle A, and Johnson LF. Drivers of sex differences in the South African adult tuberculosis incidence and mortality trends, 1990–2019. Scientific Reports 2023; 13
1990
-
[30]
The dynamics of tuberculosis transmission model with different genders
Wang S and Cao H. The dynamics of tuberculosis transmission model with different genders. Journal of Biological Dynamics 2024; 18
2024
-
[31]
Learning differential equation models from stochastic agent-based model simulations
Nardini JT, Baker RE, Simpson MJ, and Flores KB. Learning differential equation models from stochastic agent-based model simulations. Journal of the Royal Society Interface 2021; 18
2021
-
[32]
Efficient coupling of within-and between-host infectious disease dynamics
Smith CA and Ashby B. Efficient coupling of within-and between-host infectious disease dynamics. Journal of Theoretical Biology 2025
2025
-
[33]
A model of tuberculosis progression using CompuCell3D
Doran JWG, Rowlatt CF, Powathil GG, Bowness R, and Yates CA. A model of tuberculosis progression using CompuCell3D. arXiv preprint arXiv:2602.24258 2026
-
[34]
Initiation and regulation of T-cell responses in tuberculosis
Urdahl KB, Shafiani S, and Ernst JD. Initiation and regulation of T-cell responses in tuberculosis. Mucosal immunology 2011; 4
2011
-
[35]
Uncertainty and sensitivity analyses methods for agent-based mathematical models: An introductory review
Hamis S, Stratiev S, and Powathil GG. Uncertainty and sensitivity analyses methods for agent-based mathematical models: An introductory review. THE PHYSICS OF CANCER: Research Advances 2021
2021
-
[36]
When is an outbreak not an outbreak? Fit, divergent strains of Mycobacterium tuberculosis display independent evolution of drug resistance in a large London outbreak
Shorten RJ, McGregor AC, Platt S, Jenkins C, Lipman MCI, Gillespie SH, Charalambous BM, and McHugh TD. When is an outbreak not an outbreak? Fit, divergent strains of Mycobacterium tuberculosis display independent evolution of drug resistance in a large London outbreak. Journal of Antimicrobial Chemotherapy 2013; 68
2013
-
[37]
A flow cytometry method for rapidly assessing Mycobacterium tuberculosis responses to antibiotics with different modes of action
Hendon-Dunn CL, Doris KS, Thomas SR, Allnutt JC, Marriott AAN, Hatch KA, Watson RJ, Bottley G, Marsh PD, and Taylor SC. A flow cytometry method for rapidly assessing Mycobacterium tuberculosis responses to antibiotics with different modes of action. Antimicrobial agents and chemotherapy 2016; 60
2016
-
[38]
Quantitative study on the production and kinetics of mononuclear phagocytes during an acute inflammatory reaction
Van Furth R, Diesselhoff-den Dulk MMC, and Mattie H. Quantitative study on the production and kinetics of mononuclear phagocytes during an acute inflammatory reaction. The Journal of Experimental Medicine 1973; 138
1973
-
[39]
Identifying control mechanisms of granuloma formation during M
Segovia-Juarez JL, Ganguli S, and Kirschner D. Identifying control mechanisms of granuloma formation during M. tuberculosis infection using an agent-based model. Journal of Theoretical Biology 2004; 231
2004
-
[40]
Lifespans of naive, memory and effector lymphocytes
Sprent J. Lifespans of naive, memory and effector lymphocytes. Current Opinion in Immunology 1993; 5 32
1993
-
[41]
The global burden of latent tuberculosis infection: a re-estimation using mathematical modelling
Houben RMGJ and Dodd PJ. The global burden of latent tuberculosis infection: a re-estimation using mathematical modelling. PLOS Medicine 2016; 13
2016
-
[42]
Latent tuberculosis infection: An overview
Kiazyk S and Ball TB. Latent tuberculosis infection: An overview. Canada Communicable Disease Report 2017; 43
2017
-
[43]
Ending tuberculosis by 2030: can we do it? International Journal of Tuberculosis and Lung Disease 2016; 20
Suthar AB, Zachariah R, and Harries AD. Ending tuberculosis by 2030: can we do it? International Journal of Tuberculosis and Lung Disease 2016; 20
2030
-
[44]
Approximate accelerated stochastic simulation of chemically reacting systems
Gillespie DT. Approximate accelerated stochastic simulation of chemically reacting systems. The Journal of Chemical Physics 2001; 115
2001
-
[45]
Doran JWG, Mujuni D, Gallagher K, Yates CA, and Bowness R. An agent-based modelling approach to investigate the impact of gender on tuberculosis transmission in Uganda. arXiv preprint arXiv:2601.09813 2026
-
[46]
Risk factors for pulmonary cavitation in tuberculosis patients from China
Zhang L, Pang Y, Yu X, Wang Y, Lu J, Gao M, Huang H, and Zhao Y. Risk factors for pulmonary cavitation in tuberculosis patients from China. Emerging microbes & infections 2016; 5
2016
-
[47]
Systematic review and meta-analysis of sex differences in social contact patterns and implications for tuberculosis transmission and control
Horton KC, Hoey AL, B´ eraud G, Corbett EL, and White RG. Systematic review and meta-analysis of sex differences in social contact patterns and implications for tuberculosis transmission and control. Emerging Infectious Diseases 2020; 26
2020
-
[48]
Risk factors for infectiousness of patients with tuberculosis: a systematic review and meta-analysis
Melsew YA, Doan TN, Gambhir M, Cheng AC, McBryde E, and Trauer JM. Risk factors for infectiousness of patients with tuberculosis: a systematic review and meta-analysis. Epidemiology & Infection 2018; 146
2018
-
[49]
Individual-based modeling of tuberculosis in a user-friendly interface: understanding the epidemiological role of population heterogeneity in a city
Prats C, Monta˜ nola-Sales C, Gilabert-Navarro JF, Valls J, Casanovas-Garcia J, Vilaplana C, Cardona PJ, and L´ opez D. Individual-based modeling of tuberculosis in a user-friendly interface: understanding the epidemiological role of population heterogeneity in a city. Frontiers in Microbiology 2016; 6
2016
-
[50]
Exploratory data analysis
Tukey JW. Exploratory data analysis. Vol. 2. Springer, 1977
1977
-
[51]
Factors associated with DELAY in diagnosis among tuberculosis patients in Hohoe Municipality, Ghana
Osei E, Akweongo P, and Binka F. Factors associated with DELAY in diagnosis among tuberculosis patients in Hohoe Municipality, Ghana. BMC public health 2015; 15
2015
-
[52]
Total delay in treatment among smear positive pulmonary tuberculosis patients in five primary health centers, southern Ethiopia: a cross sectional study
Asefa A and Teshome W. Total delay in treatment among smear positive pulmonary tuberculosis patients in five primary health centers, southern Ethiopia: a cross sectional study. PloS one 2014; 9
2014
-
[53]
Survival of people with untreated TB: effects of time, geography and setting
Rodriguez CA, Leavitt SV, Bouton TC, Horsburgh CR, Zur Wiesch PA, Nichols B, Jenkins HE, and White LF. Survival of people with untreated TB: effects of time, geography and setting. The International Journal of Tuberculosis and Lung Disease 2023; 27
2023
-
[54]
Treatment of tuberculosis: guidelines
World Health Organization STI(HO. Treatment of tuberculosis: guidelines. World Health Organization, 2010
2010
-
[55]
Association between tuberculosis in men and social network structure in Kampala, Uganda
Miller PB, Zalwango S, Galiwango R, Kakaire R, Sekandi J, Steinbaum L, Drake JM, Whalen CC, and Kiwanuka N. Association between tuberculosis in men and social network structure in Kampala, Uganda. BMC Infectious Diseases 2021; 21
2021
-
[56]
Learning surrogate equations for the analysis of an agent-based cancer model
Burrage K, Burrage PM, Kreikemeyer JN, Uhrmacher AM, and Weerasinghe HN. Learning surrogate equations for the analysis of an agent-based cancer model. Frontiers in Applied Mathematics and Statistics 2025; 11
2025
-
[57]
Synergy between individual TNF-dependent functions determines granuloma performance for controlling Mycobacterium tuberculosis infection
Ray JCJ, Flynn JL, and Kirschner DE. Synergy between individual TNF-dependent functions determines granuloma performance for controlling Mycobacterium tuberculosis infection. The Journal of Immunology 2009; 182 33 Appendix A Comparison of regression methods We used linear regression to infer the learnt equations for the seven cell types. We considered thr...
2009
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.