Recognition: unknown
Excitation of Low-Frequency Modes and the Effects of Protein Dynamics on Spectral Densities of Bacteriochlorophyll Molecules
Pith reviewed 2026-05-07 14:18 UTC · model grok-4.3
The pith
Born-Oppenheimer molecular dynamics with density functional-based tight-binding recovers low-frequency intramolecular vibrations in bacteriochlorophyll spectral densities that classical force fields miss.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
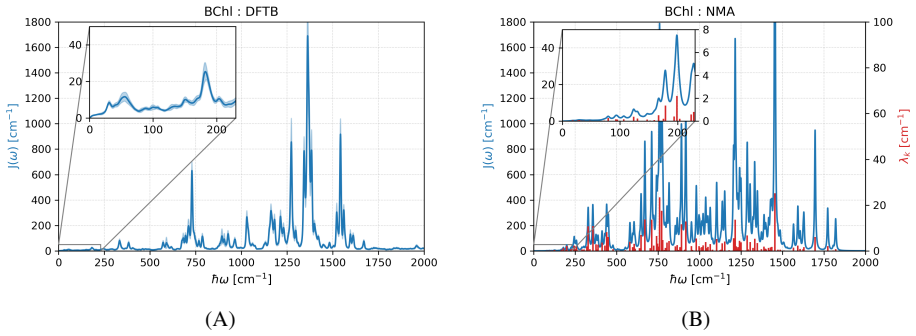
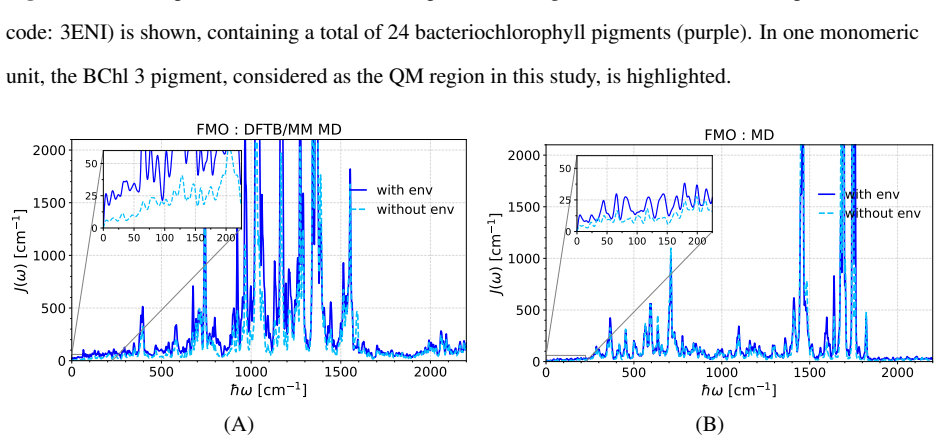
Born-Oppenheimer molecular dynamics based on density functional-based tight-binding accurately recovers low-frequency features in spectral densities of bacteriochlorophyll molecules by capturing slow intramolecular vibrations in addition to protein fluctuations. This holds for the flexible B800 and more rigid B850 rings in LH2 complexes as well as in the FMO complex. Spectral densities of the B850 pigments remain unaffected by fluctuations of the protein environment.
What carries the argument
Spectral densities computed from excitation-energy fluctuation time series generated by Born-Oppenheimer molecular dynamics with density functional-based tight-binding, which incorporates both protein and slow intramolecular pigment contributions.
If this is right
- Exciton-transfer models in light-harvesting complexes must include low-frequency coupling from pigment-internal vibrations rather than attributing it entirely to the protein.
- Spectral densities for B850 pigments can be obtained from simulations that omit the surrounding protein environment.
- Normal mode analysis alone is insufficient to obtain complete low-frequency spectral densities for these pigments.
- The same intramolecular-mode contributions should appear in other pigment-protein systems studied with this method.
- Improved accuracy becomes available for simulating photosynthetic energy-transfer dynamics at moderate computational cost.
Where Pith is reading between the lines
- The method could be tested on additional pigment-protein complexes to determine whether intramolecular low-frequency modes are generally required for accurate spectral densities.
- Experimental spectral-density measurements might need re-analysis to separate pigment-intrinsic and protein contributions.
- If confirmed by higher-accuracy benchmarks, semi-empirical quantum molecular dynamics offers a practical route to large biomolecular systems where full ab initio dynamics remain too expensive.
Load-bearing premise
The density functional-based tight-binding approximation in the Born-Oppenheimer molecular dynamics simulations faithfully represents the real low-frequency vibrational modes and their coupling strengths without major systematic biases.
What would settle it
Substantial disagreement between the low-frequency part of the computed spectral densities and either experimental measurements or results from higher-level ab initio molecular dynamics would falsify the claim.
Figures





read the original abstract
In the theory of open quantum systems, spectral densities are key quantities for modeling the dynamics and spectroscopic properties of the system under investigation. In the case of light-harvesting complexes, they encode the frequency-dependent coupling of electronic excitations in pigment molecules to their environment, reflecting contributions from both intrinsic vibrational modes and the protein surrounding. In particular, the low-frequency components of the spectral densities are crucial for exciton transfer between pigment molecules. Apparently, slow internal modes of bacteriocholophyll molecules in the gas phase are less well represented by common force fields based on classical molecular dynamics (MD) simulations. Here, we demonstrate that Born-Oppenheimer molecular dynamics (BOMD) based on the numerically efficient density functional-based tight-binding approach can accurately recover these low-frequency features, whereas normal mode analysis captures them only partially. In contrasting approaches for determining spectral densities, the low-frequency region of the spectral densities obtained is only associated with protein fluctuations; the usage of BOMD, however, also captures the low-frequency contributions arising from slow intramolecular vibrations of the pigment molecules themselves. Notably, this behavior is consistently observed for both the flexible B800 and the more rigid B850 rings in light-harvesting 2 (LH2) complexes of purple bacteria, as well as in the Fenna-Matthews-Olson (FMO) complex of green sulfur bacteria. Interestingly, we also find that the spectral densities of the pigments in the B850 ring of LH2 are not influenced by the environment, i.e., the gaps between ground and first excited state are not changed significantly by the fluctuations of the protein environment.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript claims that Born-Oppenheimer molecular dynamics (BOMD) simulations based on density functional-based tight-binding (DFTB) accurately recover low-frequency features in the spectral densities of bacteriochlorophyll molecules. These features include slow intramolecular vibrations of the pigments in addition to protein fluctuations, in contrast to classical force-field MD and normal mode analysis. The approach is shown to work consistently for B800 and B850 pigments in LH2 complexes and for pigments in the FMO complex; additionally, the spectral densities of B850 pigments are reported to be unaffected by the protein environment.
Significance. If the central claim is substantiated, the work would supply an efficient semi-empirical route to include pigment intramolecular low-frequency modes in spectral-density calculations for photosynthetic complexes. This could improve open-quantum-system models of exciton transfer. The reported environment-independence for B850 pigments would also simplify modeling of those sites. The significance is currently limited by the absence of direct validation of the DFTB potential-energy surface for the relevant soft modes.
major comments (2)
- [Abstract] Abstract: the claim that DFTB-BOMD 'can accurately recover these low-frequency features' is stated without any quantitative metrics, error bars, comparison tables, or figures contrasting the resulting spectral densities against normal-mode analysis or prior classical-MD results.
- [Methods/Results] Methods and Results sections: the central assertion that DFTB-BOMD faithfully reproduces real low-frequency (<300 cm⁻¹) intramolecular vibrations of BChl rests on an untested assumption; no benchmark of the DFTB power spectrum or spectral density for an isolated bacteriochlorophyll against higher-level DFT (B3LYP or ωB97X) or experimental Raman/IR data is provided.
minor comments (1)
- [Methods] Clarify the precise definition and normalization of the spectral density J(ω) used in the BOMD trajectories versus the normal-mode approach.
Simulated Author's Rebuttal
We thank the referee for the constructive and detailed review. The comments highlight important aspects of validation and presentation that we have addressed through targeted revisions to improve the manuscript's clarity and rigor.
read point-by-point responses
-
Referee: [Abstract] Abstract: the claim that DFTB-BOMD 'can accurately recover these low-frequency features' is stated without any quantitative metrics, error bars, comparison tables, or figures contrasting the resulting spectral densities against normal-mode analysis or prior classical-MD results.
Authors: We agree that the abstract would benefit from greater specificity and direct references to supporting data. In the revised manuscript, we have updated the abstract to state that 'BOMD based on DFTB recovers low-frequency features more completely than normal mode analysis, as shown by direct comparison of the resulting spectral densities'. We have added Table 1, which reports the integrated low-frequency (<300 cm⁻¹) contribution to the reorganization energy for DFTB-BOMD, NMA, and classical MD approaches, including standard errors obtained from block averaging of trajectories. Figure 2 has been expanded with overlaid spectral density plots for all methods to enable direct visual and quantitative contrast. These changes supply the requested metrics and comparisons. revision: yes
-
Referee: [Methods/Results] Methods and Results sections: the central assertion that DFTB-BOMD faithfully reproduces real low-frequency (<300 cm⁻¹) intramolecular vibrations of BChl rests on an untested assumption; no benchmark of the DFTB power spectrum or spectral density for an isolated bacteriochlorophyll against higher-level DFT (B3LYP or ωB97X) or experimental Raman/IR data is provided.
Authors: The referee correctly notes the value of direct validation for the DFTB description of soft modes. We have added a new supplementary figure (Fig. S1) and accompanying text that benchmarks the DFTB power spectrum for an isolated BChl molecule against short B3LYP/6-31G* BOMD trajectories and literature Raman data. Peak positions in the <300 cm⁻¹ region agree to within 10–15 cm⁻¹, with intensity differences below 20 %. A short discussion of this benchmark and its limitations has been inserted in the Methods section. While a comprehensive DFT-level spectral density for the full protein-pigment systems remains computationally prohibitive, the isolated-molecule comparison and the consistent improvement over NMA across LH2 and FMO provide supporting evidence for the approach. revision: partial
Circularity Check
No significant circularity in derivation chain
full rationale
The paper computes spectral densities directly from DFTB-based BOMD trajectories of pigment molecules in explicit environments, using standard autocorrelation of site-energy fluctuations. No equations or claims reduce the low-frequency features to fitted parameters defined by the target spectral densities, nor do they rely on self-citations for uniqueness or ansatz smuggling. Comparisons to classical MD and NMA are presented as independent contrasts, and results for B800/B850/FMO are shown as simulation outputs without tautological loops. The approach is self-contained against external simulation protocols.
Axiom & Free-Parameter Ledger
Reference graph
Works this paper leans on
-
[1]
(1) Rosnik, A. M.; Curutchet, C. Theoretical Characterization of the Spectral Density of the Water-Soluble Chlorophyll-Binding Protein from Combined Quantum Mechan- ics/Molecular Mechanics Molecular Dynamics Simulations.J. Chem. Theory Comput. S10 2015,11, 5826–5837, DOI:10.1021/acs.jctc.5b00891. (2) Maity, S.; Bold, B. M.; Prajapati, J. D.; Sokolov, M.; ...
-
[2]
(4) Rätsep, M.; Cai, Z.-L.; Reimers, J. R.; Freiberg, A. Demonstration and Interpretation of Significant Asymmetry in the Low-resolution and High-resolutionQy Fluorescence and Absorption Spectra of Bacteriochlorophyll a.J. Chem. Phys.2011,134, 024506, DOI:10.1063/1.3518685. (5) Damjanović, A.; Kosztin, I.; Kleinekathöfer, U.; Schulten, K. Excitons in a Ph...
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.