Recognition: unknown
Parsimonious computational inference protocol for Boolean networks: Application to osteogenesis
Pith reviewed 2026-05-07 12:34 UTC · model grok-4.3
The pith
A pruning protocol narrows 51,138 Boolean network candidates for osteogenesis down to six biologically consistent models.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
The paper establishes an incremental pruning protocol that, by exhaustive exploration of local function substitutions followed by filters based on interaction digraph similarity, attraction basin topology, trajectorial isomorphism, and minimization of dynamical instability and frustration, reduces a syntactic search space of 51,138 candidate Boolean networks for the osteogenesis regulation network to a robust family of six parsimonious models that preserve known biologically relevant attractors and eliminate spurious ones.
What carries the argument
The incremental pruning protocol that successively applies structural digraph similarity, basin topological organization, trajectorial isomorphism, and minimization of dynamical instability and frustration to filter candidate models.
If this is right
- The protocol produces models whose asymptotic behaviors align strictly with observed osteogenesis dynamics.
- Only six networks survive as parsimonious candidates compatible with all imposed biological constraints.
- The same filtering sequence can be reused on other multi-node genetic control networks.
- The resulting models contain fewer extraneous regulatory interactions than the initial candidate set.
- Dynamical instability and frustration are measurably lower in the retained models than in the unpruned space.
Where Pith is reading between the lines
- The six models could serve as starting points for targeted experimental validation of specific regulatory edges in bone differentiation.
- If the pruning criteria prove portable, the approach could reduce the computational cost of Boolean-network inference across other developmental or disease-related gene networks.
- One could test whether adding quantitative expression thresholds as an extra filter further shrinks the surviving model family.
- The protocol's emphasis on attractor preservation suggests it might help distinguish core regulatory motifs from peripheral ones in larger networks.
Load-bearing premise
The selected pruning criteria correctly separate biologically viable models from spurious ones without discarding valid alternatives or retaining invalid ones.
What would settle it
Experimental tests of gene-expression trajectories under perturbations that the six models predict differently from one another but that current biological data do not yet resolve.
Figures
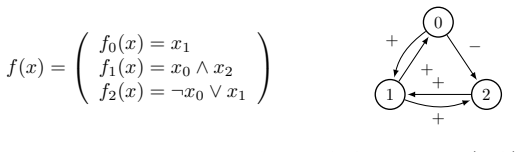



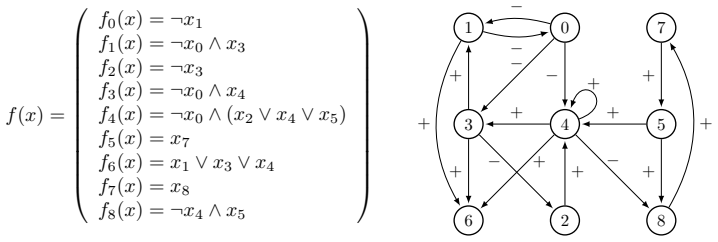
read the original abstract
Boolean networks are powerful mathematical tools for modeling the qualitative dynamics of genetic regulation. Yet inferred models often generate spurious attractors that lack biological viability. In this paper, we propose a parsimonious computational framework to systematically refine Boolean network models by eliminating these non-biological asymptotic behaviors while strictly preserving known, biologically relevant attractors. Through an exhaustive exploration of local function substitutions, we generate a comprehensive set of candidate models. To identify the most biologically consistent networks, we implement an incremental pruning protocol that filters candidates based on structural interaction digraph similarity, attraction basin topological organization, trajectorial isomorphism, and the minimization of dynamical instability and frustration. We apply this methodology to a 9-node genetic control model of the osteogenesis regulation network. Our protocol effectively evaluates a syntactic search space of 51,138 potential networks, ultimately narrowing them down to a robust family of 6 parsimonious models that are fully compatible with current biological knowledge.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript presents a parsimonious computational protocol for refining Boolean network models of gene regulation. It exhaustively generates 51,138 candidate networks for a 9-node osteogenesis regulatory graph via local function substitutions, then applies an incremental pruning protocol based on structural digraph similarity, attraction basin topology, trajectorial isomorphism, dynamical instability, and frustration to retain a family of 6 models asserted to preserve known biological attractors while eliminating spurious ones.
Significance. If the protocol's criteria reliably separate biologically viable models from spurious ones, the work would supply a systematic, reproducible method for addressing a common limitation of Boolean network inference—unwanted attractors—while remaining grounded in external biological knowledge. The explicit enumeration of the full syntactic space and step-wise filtering in the osteogenesis case provides a concrete, falsifiable demonstration that could be adapted to other small-to-medium gene regulatory networks.
major comments (2)
- [Results (osteogenesis application)] Results section on the final 6 models: compatibility with biological knowledge is asserted via matching of literature-reported fixed points and limit cycles, yet no quantitative validation (e.g., basin-size error, perturbation robustness, or comparison to time-series data) is supplied to confirm the retained models reproduce known osteogenesis dynamics without introducing new non-biological behaviors.
- [Methods (incremental pruning protocol)] Methods (pruning criteria definition): the claim that the five filtering steps correctly distinguish viable from spurious models rests on the untested assumption that the chosen thresholds and similarity measures do not over-prune valid alternatives; no synthetic benchmark or cross-validation on held-out biological constraints is presented, which is load-bearing for the central reduction claim.
minor comments (2)
- [Abstract] Abstract: the reduction from 51,138 to 6 models is stated without indicating the specific validation performed on the retained models, which would help readers assess the strength of the compatibility claim.
- [Figures] Figure legends: ensure all panels displaying interaction digraphs and attractor lists for the final models include explicit node labels, threshold values, and a statement of which literature sources define the target attractors.
Simulated Author's Rebuttal
We thank the referee for the constructive report and the opportunity to clarify our work. We address each major comment point-by-point below, committing to revisions where they strengthen the manuscript without altering its core claims or scope.
read point-by-point responses
-
Referee: Results section on the final 6 models: compatibility with biological knowledge is asserted via matching of literature-reported fixed points and limit cycles, yet no quantitative validation (e.g., basin-size error, perturbation robustness, or comparison to time-series data) is supplied to confirm the retained models reproduce known osteogenesis dynamics without introducing new non-biological behaviors.
Authors: The validation in the manuscript rests on exhaustive enumeration followed by filters that explicitly preserve the literature-reported attractors while discarding models that introduce additional attractors or violate structural/topological/dynamical consistency with the known osteogenesis network. This approach directly addresses the common problem of spurious attractors in Boolean network inference. We agree that quantitative supplements would be beneficial. In revision we will add a supplementary table comparing attraction basin sizes of the six retained models against the unpruned network and will include a brief perturbation analysis (single-node flips) to quantify robustness. Direct comparison to time-series data is not feasible, as the model is constructed from qualitative regulatory literature rather than quantitative kinetic measurements; we will state this limitation explicitly in the revised discussion. revision: partial
-
Referee: Methods (pruning criteria definition): the claim that the five filtering steps correctly distinguish viable from spurious models rests on the untested assumption that the chosen thresholds and similarity measures do not over-prune valid alternatives; no synthetic benchmark or cross-validation on held-out biological constraints is presented, which is load-bearing for the central reduction claim.
Authors: The thresholds and similarity measures were set to the minimal values that still guarantee retention of all literature-reported attractors after each filtering stage, as verified by the exhaustive search. The protocol therefore provides a falsifiable, reproducible demonstration on a concrete biological example rather than a general proof of optimality. We acknowledge that synthetic benchmarks or cross-validation on held-out constraints would be required to establish broader validity of the specific numerical thresholds. In the revised manuscript we will expand the Methods section with explicit justification for each threshold choice and add a dedicated limitations paragraph noting the absence of synthetic benchmarks and the consequent need for future testing on additional networks. revision: partial
Circularity Check
No significant circularity identified
full rationale
The paper's protocol generates candidate Boolean networks via exhaustive substitution rules on a fixed 9-node graph and then applies sequential external filters (digraph similarity, basin topology, trajectorial isomorphism, instability, frustration) that are evaluated against independently known biological attractors from the literature. No derived quantity is obtained by fitting parameters to the target result, no equation reduces to its own input by definition, and no load-bearing premise rests on a self-citation chain. The final selection of six models is presented as an explicit enumeration that preserves reported fixed points and cycles while discarding others, making the derivation self-contained against external benchmarks.
Axiom & Free-Parameter Ledger
Reference graph
Works this paper leans on
-
[1]
Costamagna, B
E. Costamagna, B. García, and P. Santisteban. The functional interaction between the paired domain transcription factor Pax8 and Smad3 is in- volved in transforming growth factor-beta repression of the sodium/iodide symporter gene.J. Biol. Chem., 279:3439–3446, 2004
2004
-
[2]
Demongeot, A
J. Demongeot, A. K. Diallo, A. Hazgui, H. Espinoza, and M. Montalva- Medel. Boolean networks with classic and new updating modes applied to genetic regulation in some familial diseases.Int. J. Mol. Sci., 26:11976, 2025
2025
-
[3]
Demongeot, M
J. Demongeot, M. Jelassi, H. Hazgui, S. Ben Miled, N. Bellamine Ben Saoud, et al. Biological networks entropies: Examples in neural mem- ory networks, genetic regulation networks and social epidemic networks. Entropy, 20:36, 2018
2018
-
[4]
Demongeot, M
J. Demongeot, M. Noual, and S. Sené. Combinatorics of Boolean automata circuits dynamics.Discrete Appl. Math., 160:398–415, 2012
2012
-
[5]
Goles and S
E. Goles and S. Martínez.Neural and automata networks: dynamical be- havior and applications. Kluwer Academic Publishers, 1990
1990
-
[6]
C. S. Jayasena, T. Ohyama, and N. Segil. Notch signaling augments the canonical Wnt pathway to specify the size of the otic placode.Development, 135:2251–2261, 2008
2008
-
[7]
H. C. Kang, M. Ohmori, N. Harii, T. Endo, and T. Onaya. Pax-8 is essential for regulation of the thyroglobulin gene by transforming growth factor-beta1.Endocrinology, 142:267–275, 2001
2001
-
[8]
Metabolicstabilityandepigenesisinrandomlyconstructed genetic nets.J
S.A.Kauffman. Metabolicstabilityandepigenesisinrandomlyconstructed genetic nets.J. Theor. Biol., 22:437–467, 1969
1969
-
[9]
W. S. McCulloch and W. Pitts. A logical calculus of the ideas immanent in nervous activity.J. Math. Biophys., 5:115–133, 1943
1943
-
[10]
F. Ni, J. Li, Q. Yin, Y. Chen, Z. Shao, et al. Novel lncrna Lncmstrg.11341.25 promotes osteogenic differentiation of human bone marrow stem cells via the mir-939-5p/pax8 axis.Research (Wash D.C.), 8:0601, 2025
2025
-
[11]
Noual and S
M. Noual and S. Sené. Synchronism versus asynchronism in monotonic Boolean automata networks.Nat. Comput., 17:393–402, 2018. 15
2018
-
[12]
S. A. Patel, S. Hirosue, P. Rodrigues, E. Vojtasova, E. K. Richardson, et al. The renal lineage factorpax8 controls oncogenic signalling in kidney cancer.Nature, 606:999–1006, 2022
2022
-
[13]
Entropyasarobustness marker in genetic regulatory networks.Entropy, 22:260, 2020
M.Rachdi, J.Waku, H.Hazgui, andJ.Demongeot. Entropyasarobustness marker in genetic regulatory networks.Entropy, 22:260, 2020
2020
-
[14]
F. Robert. Itérations sur des ensembles finis et automates cellulaires con- tractants.Linear Algebra Appl., 29:393–412, 1980
1980
-
[15]
Robert.Discrete iterations: a metric study
F. Robert.Discrete iterations: a metric study. Springer, 1986
1986
-
[16]
R. Thomas. Boolean formalization of genetic control circuits.J. Theor. Biol., 42:563–585, 1973
1973
-
[17]
R. Thomas. On the relation between the logical structure of systems and their ability to generate multiple steady states or sustained oscillations. InNumerical methods in the study of critical phenomena, pages 180–193. Springer, 1981
1981
-
[18]
Y. Tian, Y. Xu, T. Xue, L. Chen, B. Shi, et al. Notch activation enhances mesenchymal stem cell sheet osteogenic potential by inhibition of cellular senescence.Cell Death & Disease, 8:e2595, 2017
2017
-
[19]
Toulouse
G. Toulouse. Theory of the frustration effect in spin glasses. I.Commun. Phys., 2:115–119, 1977
1977
-
[20]
Zanotti and E
S. Zanotti and E. Canalis. Notch signaling and the skeleton.Endocr. Rev., 37:223–253, 2016. 16
2016
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.