Recognition: unknown
Equilibrium and Competition in Evolutionary Dynamics
Pith reviewed 2026-05-10 14:44 UTC · model grok-4.3
The pith
Polymerization cascades reach a unique positive equilibrium above a sharp resource threshold, while replication equilibria survive only under neutral fitness.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
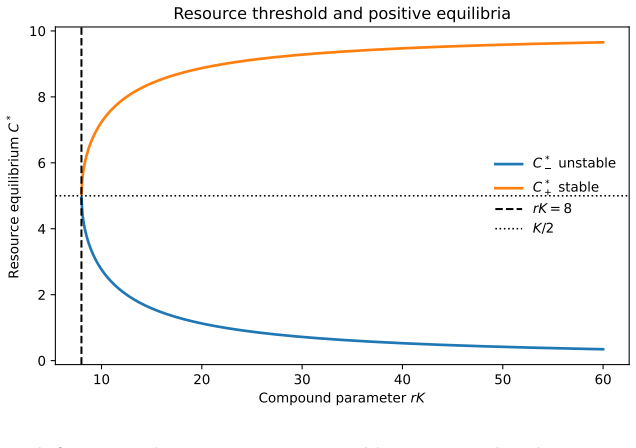
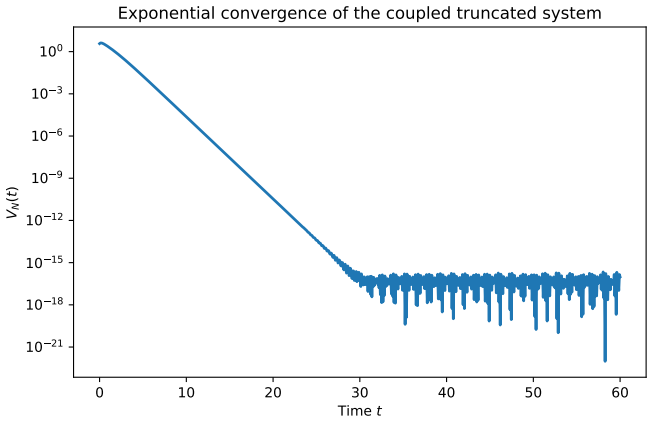
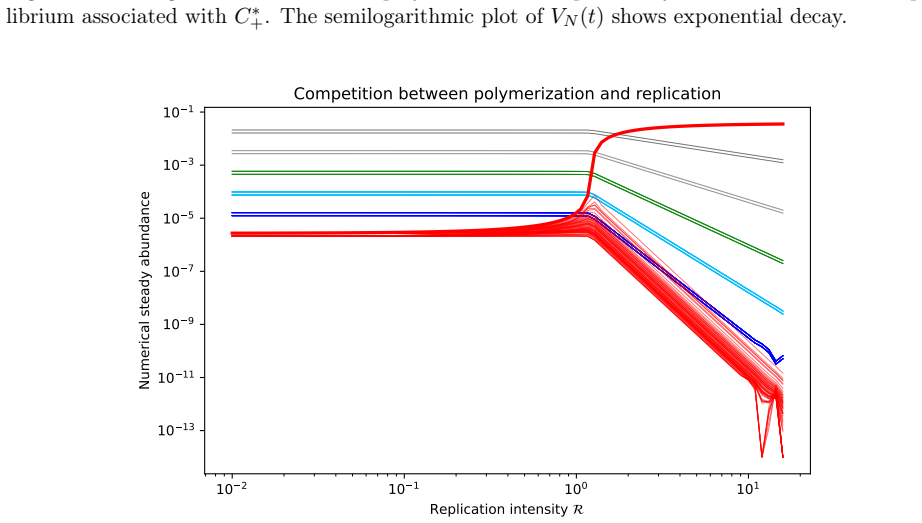
Setting fixed activated precursors, the polymerization system is globally well-posed, positive, and converges exponentially to a unique strictly positive equilibrium distribution. When the same cascades compete for a logistic resource, strictly positive equilibria exist exactly above a sharp resource threshold, with the lower branch unstable and the upper branch locally stable. For the infinite system under persistence and dominance assumptions, global exponential stability holds. Introducing a replicator term shows that the pre-replicative equilibrium continues only under neutral fitness and is removed by heterogeneous fitness.
What carries the argument
The genealogical structure of finite sequences in the open polymerization cascade, together with the three-dimensional core subsystem that governs equilibrium structure in the resource-coupled competition model.
If this is right
- Strictly positive equilibria exist exactly above the sharp resource threshold.
- The lower equilibrium branch is unstable and the upper branch is locally stable.
- Under an additional uniform decay condition, convergence is globally exponentially stable in the uniform norm.
- The pre-replicative equilibrium persists only under neutral fitness and is removed by heterogeneous fitness.
- The infinite system satisfies positivity, global existence, and a priori bounds, with global exponential stability under persistence and dominance.
Where Pith is reading between the lines
- The sharp threshold supplies a concrete resource level below which no persistent pre-replicative sequence distributions can form.
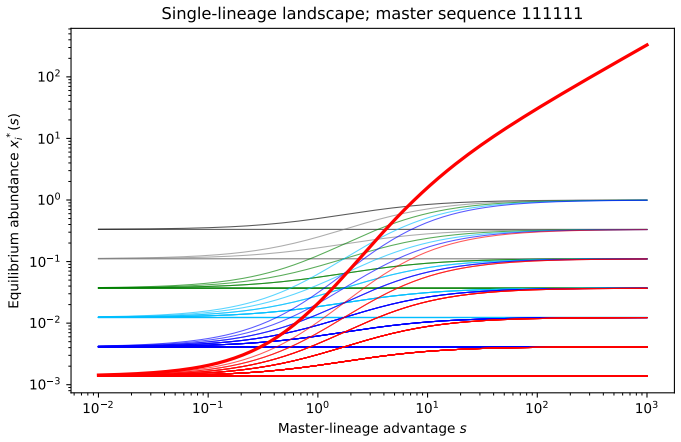
- Heterogeneous fitness removing the pre-replicative equilibrium implies that any early replicators must operate away from the neutral pre-replication state, potentially requiring additional mechanisms to sustain diversity.
Load-bearing premise
The assumption that growth follows deterministic differential equations along fixed genealogical chains with logistic resource depletion and no stochastic fluctuations or spatial structure.
What would settle it
Direct measurement of whether polymer sequence distributions reach a persistent positive steady state only when resource levels exceed the model's calculated threshold, or whether varying fitness values eliminate that steady state once replication is introduced.
Figures


read the original abstract
A fundamental problem in protobiological dynamics is to understand how chemically generated polymers can form persistent sequence distributions before the emergence of replication. We study deterministic polymer growth in which each finite sequence is followed along its genealogical structure. The system pictures an open polymerization cascade in which each polymer is produced from a unique precursor and lost by degradation and further extension. Setting fixed activated precursors, we show global well-posedness, positivity, uniqueness of a strictly positive equilibrium, and exponential convergence to an explicit steady state distribution. Under an additional uniform decay condition, this convergence becomes global exponential stability in a uniform norm. We then couple the polymerization to a shared environmental resource with logistic growth and depletion by two activated precursors. In the resulting binary polymerization competition model, the equilibrium structure is governed by a three-dimensional core subsystem. We prove that strictly positive equilibria exist exactly above a sharp resource threshold. At the threshold the equilibrium is unique, while above it two positive branches appear. The lower branch is unstable and the upper branch is locally stable. For the complete infinite system, we exhibit positivity, global componentwise existence, a priori bounds, and under persistence and dominance assumptions, global exponential stability. Finally, we introduce template directed replication through a replicator term. The pre-replicative equilibrium continues only under neutral fitness, and heterogeneous fitness removes it as an equilibrium of the replicated system.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper analyzes deterministic models of polymer growth in open cascades, proving global well-posedness, positivity, uniqueness of a strictly positive equilibrium, and exponential convergence to an explicit steady-state distribution for fixed activated precursors (with global exponential stability in uniform norm under an additional uniform decay condition). It then couples the system to logistic resource dynamics with two precursors, reducing equilibria to a 3D core and proving strictly positive equilibria exist above a sharp resource threshold (unique at threshold, with lower unstable and upper locally stable branches above it). For the full infinite system, positivity, a priori bounds, and global exponential stability are shown under persistence and dominance assumptions. Finally, template-directed replication is added, with the pre-replicative equilibrium persisting only under neutral fitness and being removed under heterogeneous fitness.
Significance. If the stability and existence results hold, the work supplies a rigorous mathematical framework for prebiotic polymer sequence distributions, resource competition, and the transition to replication, including explicit steady states and sharp thresholds. The direct analysis without fitted parameters and the reduction of the infinite competition model to a 3D core are strengths that could inform origins-of-life modeling.
major comments (3)
- [Fixed activated precursors section] In the fixed-precursor analysis, global exponential stability in the uniform norm is claimed only under an extra uniform decay condition, but no verification is given that this condition holds for general rate parameters or fitness landscapes; this assumption is load-bearing for the global stability claim and the subsequent removal of the pre-replicative equilibrium under heterogeneous fitness.
- [Binary polymerization competition model] For the coupled logistic-resource model, the infinite-dimensional system requires separate persistence and dominance assumptions to obtain global componentwise existence, a priori bounds, and global exponential stability, yet the manuscript provides no general proof or parameter ranges where these hold; if they fail for some fitness landscapes, the claimed global stability and the 3D-core reduction do not extend to the full system.
- [Template-directed replication section] The statement that heterogeneous fitness removes the pre-replicative equilibrium as an equilibrium of the replicated system is central, but the derivation in the infinite-dimensional setting with the replicator term is not shown explicitly; the reduction to the 3D core and the effect on positivity/uniqueness need to be detailed to confirm the claim.
minor comments (2)
- [Introduction] Notation for the infinite sequence space and the genealogical structure could be clarified with an explicit diagram or example for a short polymer.
- [Fixed activated precursors section] The abstract mentions 'exponential convergence to an explicit steady state distribution' but the main text should cross-reference the exact formula for this distribution.
Simulated Author's Rebuttal
We thank the referee for their careful reading and constructive comments, which have helped us identify areas for clarification. We address each major comment point by point below, with plans for targeted revisions to strengthen the presentation of the results.
read point-by-point responses
-
Referee: [Fixed activated precursors section] In the fixed-precursor analysis, global exponential stability in the uniform norm is claimed only under an extra uniform decay condition, but no verification is given that this condition holds for general rate parameters or fitness landscapes; this assumption is load-bearing for the global stability claim and the subsequent removal of the pre-replicative equilibrium under heterogeneous fitness.
Authors: We agree that the uniform decay condition is an additional hypothesis needed for global exponential stability in the uniform norm and that its verification for general parameters would strengthen the result. The manuscript presents the stability as conditional on this assumption rather than claiming it universally. In revision we will add a dedicated paragraph with sufficient conditions (e.g., uniform lower bounds on degradation rates relative to extension rates) under which the decay condition holds for bounded fitness landscapes, together with a brief verification argument. This clarifies applicability without altering the stated theorems. revision: partial
-
Referee: [Binary polymerization competition model] For the coupled logistic-resource model, the infinite-dimensional system requires separate persistence and dominance assumptions to obtain global componentwise existence, a priori bounds, and global exponential stability, yet the manuscript provides no general proof or parameter ranges where these hold; if they fail for some fitness landscapes, the claimed global stability and the 3D-core reduction do not extend to the full system.
Authors: The persistence and dominance assumptions are introduced precisely to guarantee well-posedness and stability for the infinite system, consistent with standard practice in infinite-dimensional population models. We acknowledge that explicit parameter ranges and proofs are not supplied. In the revised manuscript we will insert a new proposition stating sufficient conditions (lower bounds on resource inflow and upper bounds on fitness variation) that imply persistence and dominance, together with a short proof sketch. This will make the extension of the 3D-core reduction and global stability results more explicit. revision: yes
-
Referee: [Template-directed replication section] The statement that heterogeneous fitness removes the pre-replicative equilibrium as an equilibrium of the replicated system is central, but the derivation in the infinite-dimensional setting with the replicator term is not shown explicitly; the reduction to the 3D core and the effect on positivity/uniqueness need to be detailed to confirm the claim.
Authors: We thank the referee for highlighting the need for an explicit derivation. The manuscript outlines that the replicator term removes the pre-replicative equilibrium under heterogeneous fitness, but the infinite-dimensional substitution is only summarized. We will expand this section with the direct computation: inserting the candidate equilibrium into the augmented equations shows that the replicator contribution is nonzero unless fitness is neutral, thereby eliminating the equilibrium. We will also detail how this affects the 3D-core reduction and the preservation of positivity for the remaining equilibria. revision: yes
Circularity Check
No circularity: direct analysis of ODE system with explicit assumptions
full rationale
The paper performs standard mathematical analysis on a system of ODEs for polymer sequences and resource competition. It proves well-posedness, positivity, existence/uniqueness of equilibria, and convergence under explicitly stated additional conditions (uniform decay, persistence, dominance). These conditions are introduced as hypotheses rather than derived from the results themselves. No parameters are fitted to data, no predictions reduce to inputs by construction, and no self-citations form load-bearing chains for the core claims. The derivation chain is self-contained within the model equations and standard ODE theory.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption Polymer growth follows a deterministic open cascade where each sequence has a unique precursor
- standard math Standard local existence, positivity preservation, and continuation theorems for ODEs extend to the infinite-dimensional setting
Reference graph
Works this paper leans on
-
[1]
Schopf, J. W. (2024). Pioneers of origin of life studies--Darwin, Oparin, Haldane, Miller, Or\' o --and the oldest known records of life . Life, 14(10), 1345. https://doi.org/10.3390/life14101345
-
[2]
Approaches to the origin of life on earth
Kauffman, Stuart A (2011). Approaches to the origin of life on earth. Life 1(1), 34--48 https://doi.org/10.3390/life1010034
-
[3]
Thinking about life: the history and philosophy of biology and other sciences
Agutter, Paul S & Wheatley, Denys N (2008). Thinking about life: the history and philosophy of biology and other sciences. Springer, p. 224. https://doi.org/10.1007/978-1-4020-8866-7
-
[4]
Eigen, M. (1971). Selforganization of matter and the evolution of biological macromolecules. Naturwissenschaften, 58, 465--523. https://doi.org/10.1007/BF00623322
-
[5]
Eigen, M., & Schuster, P. (1977). A principle of natural self-organization. Part A: Emergence of the hypercycle. Naturwissenschaften, 64, 541--565. https://doi.org/10.1007/BF00450633
-
[6]
Nowak, M. A., & Ohtsuki, H. (2008). Prevolutionary dynamics and the origin of evolution. Proceedings of the National Academy of Sciences of the United States of America, 105(39), 14924--14927. https://doi.org/10.1073/pnas.0806714105
-
[7]
Manapat, M., Ohtsuki, H., Bürger, R., & Nowak, M. A. (2009). Originator dynamics. Journal of Theoretical Biology, 256(4), 586--595. https://doi.org/10.1016/j.jtbi.2008.10.006
-
[8]
Sutherland, J. D. (2017). Opinion: Studies on the origin of life --- the end of the beginning. Nature Reviews Chemistry, 1, 0012. https://doi.org/10.1038/s41570-016-0012
-
[9]
Pascal, R., Pross, A., & Sutherland, J. D. (2013). Towards an evolutionary theory of the origin of life based on kinetics and thermodynamics. Open Biology, 3, 130156. https://doi.org/10.1098/rsob.130156
-
[10]
Baaske, P., Weinert, F. M., Duhr, S., Lemke, K. H., Russell, M. J., & Braun, D. (2007). Extreme accumulation of nucleotides in simulated hydrothermal pore systems. Proceedings of the National Academy of Sciences of the United States of America, 104(22), 9346--9351. https://doi.org/10.1073/pnas.0609592104
-
[11]
Ianeselli, A., Salditt, A., Mast, C., Ercolano, B., Kufner, C. L., Scheu, B., & Braun, D. (2023). Physical non-equilibria for prebiotic nucleic acid chemistry. Nature Reviews Physics, 5, 185--195. https://doi.org/10.1038/s42254-022-00550-3
-
[12]
P., Ali, S., Yoda, I., Adam, Z
Yi, R., Tran, Q. P., Ali, S., Yoda, I., Adam, Z. R., Cleaves II, H. J., & Fahrenbach, A. C. (2020). A continuous reaction network that produces RNA precursors. Proceedings of the National Academy of Sciences of the United States of America, 117(24), 13267--13274. https://doi.org/10.1073/pnas.1922139117
-
[13]
J., Duzdevich, D., Ding, D., & Szostak, J
Zhang, S. J., Duzdevich, D., Ding, D., & Szostak, J. W. (2022). Freeze-thaw cycles enable a prebiotically plausible and continuous pathway from nucleotide activation to nonenzymatic RNA copying. Proceedings of the National Academy of Sciences of the United States of America, 119, e2116429119. https://doi.org/10.1073/pnas.2116429119
-
[14]
Yeates, J. A. M., Hilbe, C., Zwick, M., Nowak, M. A., & Lehman, N. (2016). Dynamics of prebiotic RNA reproduction illuminated by chemical game theory. Proceedings of the National Academy of Sciences of the United States of America, 113(18), 5030--5035. https://doi.org/10.1073/pnas.1525273113
-
[15]
Taylor, P. D., & Jonker, L. B. (1978). Evolutionary stable strategies and game dynamics. Mathematical Biosciences, 40, 145--156. https://doi.org/10.1016/0025-5564(78)90077-9
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.