Recognition: unknown
Optimal information transmission in a sequential model for cell division
Pith reviewed 2026-05-08 01:47 UTC · model grok-4.3
The pith
Information transmission from molecular steps to population size in dividing cells is optimal at an intermediate number of steps.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
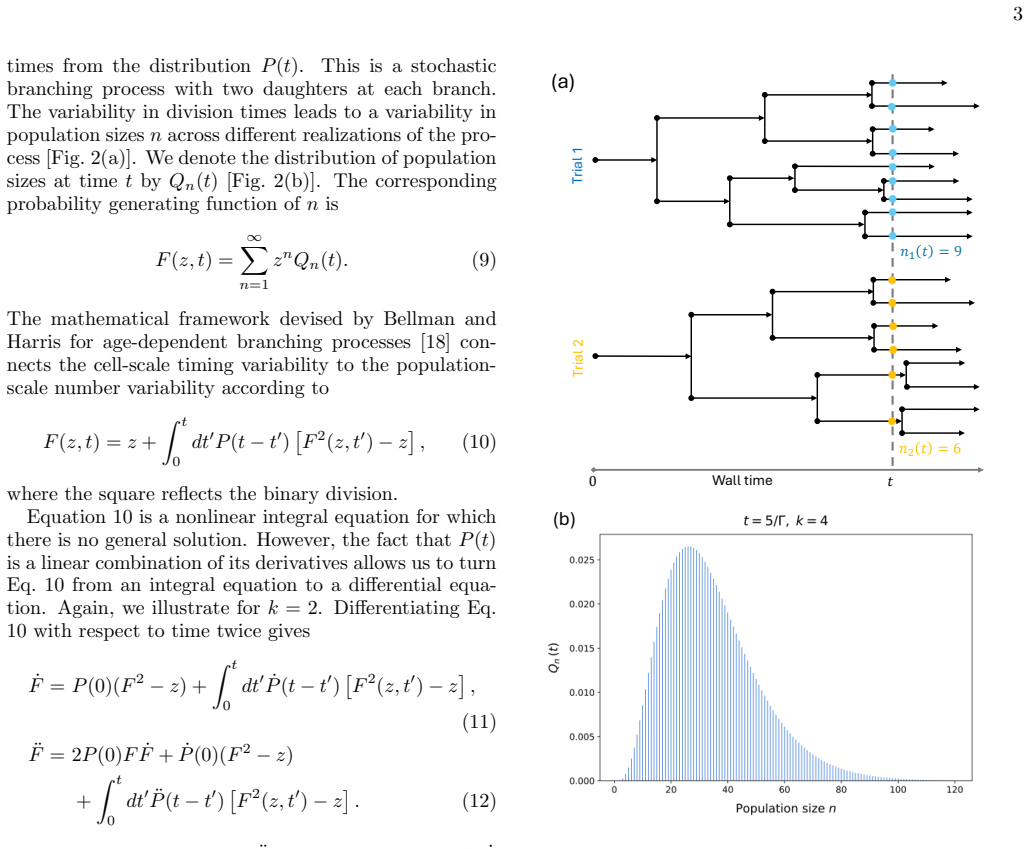
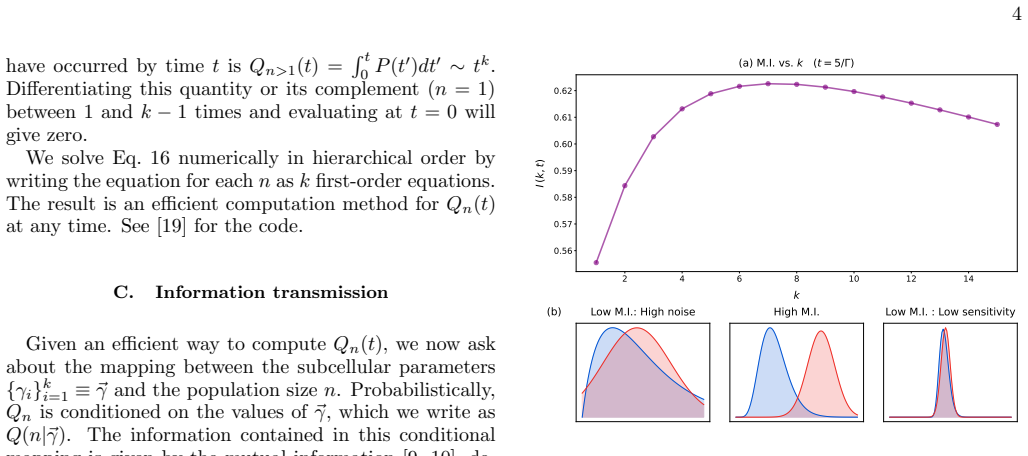
We identify a class of division time distributions, built from a series of Markovian steps, for which the population size distribution at all times is hierarchically calculable. We use this feature to characterize the amount of influence that a given reaction step has on the population size via the mutual information. We find that information transmission is optimal for a characteristic number of steps until division: too few and the population size is unpredictable; too many and any given step has vanishing influence on the population size.
What carries the argument
The class of division time distributions constructed from a finite series of Markovian steps, which permits hierarchical calculation of the population size distribution and exact evaluation of mutual information between individual steps and population size.
If this is right
- Population size becomes less predictable when cells divide after too few sequential steps.
- Individual molecular steps lose influence on population size when too many steps are required before division.
- Adaptive changes in biochemical reaction rates can optimize information flow only within a specific range of division step counts.
- Tradeoffs exist between predictability at the population scale and influence at the molecular scale.
Where Pith is reading between the lines
- Cells may have evolved division mechanisms with step numbers near this optimum to enhance adaptive responses.
- The same sequential model could apply to other biological branching processes such as lineage tracing or tumor growth.
- Experimental validation would involve varying the number of rate-limiting steps in cell cycle models and measuring population variability.
Load-bearing premise
Division times must belong to the class of distributions built from a finite series of Markovian steps so that the population size distribution remains hierarchically calculable at all times.
What would settle it
Observe whether the mutual information between a division step and population size reaches a maximum at an intermediate step count in a controlled cell population with tunable division timing.
Figures



read the original abstract
In proliferating cell populations, adaptive changes to biochemical reactions can change a cell's division time, which in turn can change the population size. However, biochemical reactions are subject to noise, and therefore the conditions for optimal information transmission from the molecular to the population scale are poorly understood. Here, we model cell proliferation as a Bellman-Harris branching process with age-dependent division times. We identify a class of division time distributions, built from a series of Markovian steps, for which the population size distribution at all times is hierarchically calculable. We use this feature to characterize the amount of influence that a given reaction step has on the population size via the mutual information. We find that information transmission is optimal for a characteristic number of steps until division: too few and the population size is unpredictable; too many and any given step has vanishing influence on the population size. Our work reveals the potential tradeoffs involved in adaptive decision making at the sub-cellular, cellular and population scales.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper models cell proliferation as a Bellman-Harris branching process with age-dependent division times restricted to phase-type distributions constructed from a finite sequence of Markovian steps. This restriction yields a hierarchically calculable population-size distribution at all times, which the authors exploit to compute the mutual information between any given reaction step and the population size. They report that this mutual information is maximized at an intermediate characteristic number of steps: too few steps render population size unpredictable, while too many steps cause any individual step to have vanishing influence.
Significance. If the optimality result holds rigorously, the work identifies a concrete tradeoff in information transmission from molecular reaction steps to population-scale observables in proliferating cells. A clear strength is the exact, non-approximate mutual-information calculation made possible by the hierarchical closure of the population-size law for the chosen class of division times; this avoids the sampling or moment-closure approximations that typically limit such analyses.
major comments (2)
- [Abstract] Abstract: the statement that the results 'reveal the potential tradeoffs involved in adaptive decision making at the sub-cellular, cellular and population scales' is not supported by any analysis outside the phase-type class; the optimality is demonstrated only for distributions that admit hierarchical population-size computation, and no argument or numerical check is given for whether an analogous optimum appears for general division-time distributions.
- [Model definition and § on hierarchical calculability] Model definition and § on hierarchical calculability: the central optimality claim rests on the exact mutual information being computable from the closed-form population-size distribution; without an explicit derivation or verification that the hierarchical property holds for arbitrary finite numbers of Markovian steps (and without additional approximations), it is impossible to confirm that the reported optimum follows rigorously rather than from an implicit parameter choice or truncation.
minor comments (2)
- Notation for the sequence of Markovian steps and the resulting phase-type density could be accompanied by a small schematic diagram to improve readability.
- The manuscript does not discuss how sensitive the location of the optimal step number is to the specific rate parameters of the exponential waiting times; a brief sensitivity check would strengthen the result.
Simulated Author's Rebuttal
We thank the referee for their careful reading and constructive comments, which help clarify the scope and rigor of our results. We respond to each major comment below and will revise the manuscript accordingly.
read point-by-point responses
-
Referee: [Abstract] Abstract: the statement that the results 'reveal the potential tradeoffs involved in adaptive decision making at the sub-cellular, cellular and population scales' is not supported by any analysis outside the phase-type class; the optimality is demonstrated only for distributions that admit hierarchical population-size computation, and no argument or numerical check is given for whether an analogous optimum appears for general division-time distributions.
Authors: We agree that the optimality result and the exact mutual-information calculation are demonstrated only within the phase-type class of division times that admit hierarchical population-size computation. The abstract's reference to 'potential tradeoffs' is intended as a forward-looking suggestion rather than a claim of generality. In revision we will rephrase the abstract to state explicitly that the reported optimum holds for this class of distributions and that analogous behavior for arbitrary division-time distributions remains an open question requiring different (typically approximate) methods. revision: yes
-
Referee: [Model definition and § on hierarchical calculability] Model definition and § on hierarchical calculability: the central optimality claim rests on the exact mutual information being computable from the closed-form population-size distribution; without an explicit derivation or verification that the hierarchical property holds for arbitrary finite numbers of Markovian steps (and without additional approximations), it is impossible to confirm that the reported optimum follows rigorously rather than from an implicit parameter choice or truncation.
Authors: The hierarchical closure arises because each phase-type distribution is realized as a linear chain of exponential waiting times; the joint state of the population can therefore be tracked by a finite system of ODEs whose dimension grows linearly with the number of steps. We will add an explicit inductive derivation in the revised manuscript showing that the population-size distribution remains exactly computable for any finite number of steps, without truncation or additional approximations. The mutual information is then obtained directly from this distribution, and the location of the maximum with respect to step number is verified by direct evaluation over a range of parameters. revision: yes
Circularity Check
No significant circularity; optimality result follows from explicit computation on self-defined phase-type model
full rationale
The paper explicitly restricts to phase-type division times (finite Markovian steps) to obtain a hierarchically closed population-size distribution, then computes mutual information between a given step and the population size. The reported optimum (characteristic number of steps) is obtained by evaluating this mutual information as a function of step count within the model; it is not presupposed by definition, fitted to data, or reduced to a self-citation. No load-bearing self-citations, smuggled ansatzes, or renamings of known results appear. The derivation is self-contained within the stated assumptions and does not equate its central claim to its inputs by construction.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption Cell proliferation can be modeled as a Bellman-Harris branching process with age-dependent division times.
- domain assumption Division times belong to the class of distributions constructed from a finite series of independent Markovian steps.
Reference graph
Works this paper leans on
-
[1]
Kussell, R
E. Kussell, R. Kishony, N. Q. Balaban, and S. Leibler, Genetics169, 1807 (2005)
2005
-
[2]
P. B. Rainey, H. J. Beaumont, G. C. Ferguson, J. Gallie, C. Kost, E. Libby, and X. X. Zhang, Microb Cell Fact10 Suppl 1, S14 (2011)
2011
-
[3]
C. D. Nadell, B. L. Bassler, and S. A. Levin, J Biol7, 27 (2008)
2008
-
[4]
L. I. Rothfield and S. S. Justice, Cell88, 581 (1997)
1997
-
[5]
Coltharp, J
C. Coltharp, J. Buss, T. M. Plumer, and J. Xiao, Proc Natl Acad Sci U S A113, E1044 (2016)
2016
-
[6]
Jonas, Current opinion in microbiology18, 54 (2014)
K. Jonas, Current opinion in microbiology18, 54 (2014)
2014
-
[7]
Raj and A
A. Raj and A. Van Oudenaarden, Cell135, 216 (2008)
2008
-
[8]
M. B. Elowitz, A. J. Levine, E. D. Siggia, and P. S. Swain, Science297, 1183 (2002)
2002
-
[9]
T. M. Cover,Elements of information theory(John Wi- ley & Sons, 1999)
1999
-
[10]
Bialek,Biophysics: searching for principles(Prince- ton University Press, 2012)
W. Bialek,Biophysics: searching for principles(Prince- ton University Press, 2012)
2012
-
[11]
C. A. Yates, M. J. Ford, and R. L. Mort, Bull Math Biol 79, 2905 (2017)
2017
-
[12]
Pugatch, Proceedings of the National Academy of Sci- ences112, 2611 (2015)
R. Pugatch, Proceedings of the National Academy of Sci- ences112, 2611 (2015)
2015
-
[13]
Amir, Physical review letters112, 208102 (2014)
A. Amir, Physical review letters112, 208102 (2014)
2014
-
[14]
Willis and K
L. Willis and K. C. Huang, Nature Reviews Microbiology 15, 606 (2017)
2017
-
[15]
Susman, M
L. Susman, M. Kohram, H. Vashistha, J. T. Nechleba, H. Salman, and N. Brenner, Proceedings of the National Academy of Sciences115, E5679 (2018)
2018
-
[16]
ElGamel, H
M. ElGamel, H. Vashistha, H. Salman, and A. Mugler, Physical Review E108, L032401 (2023)
2023
-
[17]
ElGamel and A
M. ElGamel and A. Mugler, Physical Review Letters 132, 098403 (2024)
2024
-
[18]
Bellman and T
R. Bellman and T. E. Harris, Proceedings of the National Academy of Sciences34, 601 (1948). [19]https://github.com/krishnapr0/oitsmcd
1948
-
[19]
Tkaˇ cik, A
G. Tkaˇ cik, A. M. Walczak, and W. Bialek, Physical Re- view E—Statistical, Nonlinear, and Soft Matter Physics 80, 031920 (2009)
2009
-
[20]
Micali, J
G. Micali, J. Grilli, M. Osella, and M. Cosentino Lago- marsino, Sci Adv4, eaau3324 (2018)
2018
-
[21]
Iyer-Biswas, C
S. Iyer-Biswas, C. S. Wright, J. T. Henry, K. Lo, S. Burov, Y. H. Lin, G. E. Crooks, S. Crosson, A. R. Dinner, and N. F. Scherer, Proceedings of the National Academy of Sciences of the United States of America 111, 15912 (2014)
2014
-
[22]
D. P. Haeusser and P. A. Levin, Curr Opin Microbiol11, 94 (2008)
2008
-
[23]
Colin, G
A. Colin, G. Micali, L. Faure, M. C. Lagomarsino, and S. van Teeffelen, Elife10(2021)
2021
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.