Recognition: unknown
The Incommensurability Principle in Biological Transport
Pith reviewed 2026-05-08 01:41 UTC · model grok-4.3
The pith
The conserved branching exponents in biological vascular networks arise necessarily from the incommensurability of metabolic and wave-reflection optimization costs.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
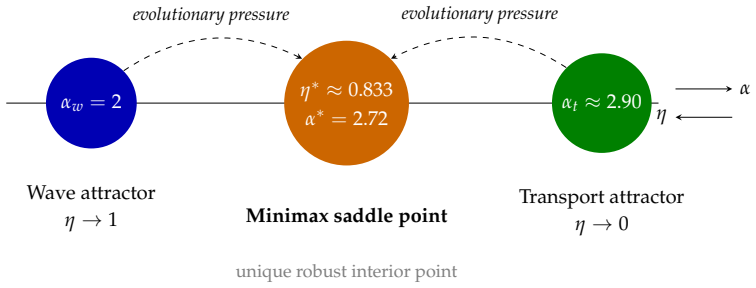
The Incommensurability Principle establishes that local optimization of extensive metabolic costs together with dimensionless wave-reflection penalties requires a coupling parameter varying by orders of magnitude across the hierarchy. The unique dimensionless cost functional consistent with scale invariance and thermodynamic linearity is the fractional metabolic excess. The minimax duty cycle η* is therefore an exact invariant of the allometric class A(G,p,α_w), orthogonal to absolute metabolic scales, and emerges as the unique attractor for networks optimizing physically incommensurable costs, with prior single-mechanism results appearing as degenerate boundary cases.
What carries the argument
The minimax duty cycle η* of the allometric class A(G,p,α_w), which carries the invariance and acts as the attractor when metabolic and wave costs are incommensurable.
If this is right
- Developmental stability of vascular architecture holds without scale-dependent tuning parameters.
- Single-mechanism optimization models appear as limiting cases of the general incommensurable-cost framework.
- The same exponents are predicted for any mammalian species whose networks belong to the allometric class.
- Changes in absolute metabolic demand do not alter the duty-cycle invariant or the resulting branching.
Where Pith is reading between the lines
- The same incommensurability logic could apply to other biological transport systems such as plant xylem or neural vasculature.
- Perturbations that alter the balance between metabolic and reflection costs should leave the exponent unchanged while shifting the duty cycle predictably.
- Evolutionary trajectories may be constrained to maintain the invariant even when absolute scales change rapidly.
- Direct measurement of duty cycles in differently sized animals would test whether the attractor value is reached independently of size.
Load-bearing premise
That the fractional metabolic excess is the unique dimensionless cost functional consistent with scale invariance and thermodynamic linearity, while alternative penalties fail empirical checks.
What would settle it
Observation of branching exponents that vary systematically with body size or metabolic rate across a wide range of species, or discovery of networks that achieve the same exponents while optimizing under a different cost measure.
Figures
read the original abstract
Biological vascular networks exhibit branching exponents ($\alpha^* \approx 2.72$) conserved across developmental stages and observed in multiple mammalian species [Kassab et al. (1993), Zamir (1999)], despite vast metabolic and anatomical variation. We prove this universality is a mathematical necessity arising from the physical incommensurability of optimization constraints. We establish three theorems. (1) No-Go Theorem: Local optimization combining extensive metabolic costs with dimensionless wave-reflection penalties requires a coupling parameter varying by $10^2$--$10^3$ across the hierarchy, precluding universal exponents. (2) Metabolic Gauge Invariance: The unique dimensionless cost functional consistent with scale invariance and thermodynamic linearity is the fractional metabolic excess; alternative penalties (logarithmic measures) fail empirical validation. (3) Architectural Invariance: The minimax duty cycle $\eta^*$ is an exact invariant of the allometric class $\mathcal{A}(G,p,\alpha_w)$, orthogonal to absolute metabolic scales -- explaining developmental stability. The minimax emerges as the unique attractor for networks optimizing physically incommensurable costs, unifying previous single-mechanism results as degenerate boundary cases.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper claims that the conserved branching exponent α* ≈ 2.72 in mammalian vascular networks is a mathematical necessity arising from the physical incommensurability of optimization constraints. It establishes three theorems: (1) a No-Go Theorem showing that local optimization of extensive metabolic costs plus dimensionless wave-reflection penalties requires a hierarchy-dependent coupling parameter, precluding universal exponents; (2) Metabolic Gauge Invariance, asserting that the fractional metabolic excess is the unique dimensionless cost functional consistent with scale invariance and thermodynamic linearity (with alternatives like logarithmic measures failing empirical validation); and (3) Architectural Invariance, showing that the minimax duty cycle η* is an exact invariant of the allometric class A(G,p,α_w), serving as the unique attractor for networks optimizing incommensurable costs and explaining developmental stability while unifying prior single-mechanism results as boundary cases.
Significance. If the derivations hold, the result would provide a parameter-free, unifying mathematical foundation for allometric scaling exponents across species and developmental stages, reducing previous models (e.g., Murray's law or wave-reflection optimizations) to degenerate limits. The identification of an exact invariant orthogonal to absolute metabolic scales offers a potential explanation for observed stability without invoking additional biological mechanisms.
major comments (3)
- [Theorem 2] Theorem 2: The assertion that the fractional metabolic excess is the unique dimensionless cost functional consistent with scale invariance and thermodynamic linearity requires an exhaustive characterization of the admissible functional space. The manuscript must explicitly derive why alternatives (e.g., other linear combinations or scale-invariant penalties preserving thermodynamic linearity) are excluded, rather than relying on empirical validation alone; without this, the uniqueness step is not demonstrated and the attractor claim in Theorem 3 does not follow as a mathematical necessity.
- [Theorem 3] Theorem 3: The claim that η* is the unique minimax attractor for the allometric class A(G,p,α_w) rests on the uniqueness established in Theorem 2. If other admissible functionals exist that remain dimensionless and linear, the incommensurability argument no longer forces a single attractor, undermining the universality of α* ≈ 2.72 as a necessity rather than a possible outcome.
- [Theorem 1] No-Go Theorem (Theorem 1): The demonstration that local optimization requires a coupling parameter varying by 10^2–10^3 across the hierarchy must be shown to be independent of the specific choice of cost functional; otherwise the no-go result may be an artifact of the particular penalties chosen rather than a general consequence of incommensurability.
minor comments (2)
- The abstract and theorems reference specific numerical values (α* ≈ 2.72, coupling variation 10^2–10^3) without citing the exact empirical sources or providing the fitting procedure used to obtain them.
- Notation for the allometric class A(G,p,α_w) and the minimax duty cycle η* should be defined explicitly at first use, including the meaning of each parameter.
Simulated Author's Rebuttal
We thank the referee for the thorough review and valuable feedback on our manuscript. The comments highlight important points regarding the rigor of our uniqueness claims and the generality of the no-go result. We address each major comment below and will incorporate clarifications and expansions in a revised version to strengthen the mathematical derivations.
read point-by-point responses
-
Referee: [Theorem 2] Theorem 2: The assertion that the fractional metabolic excess is the unique dimensionless cost functional consistent with scale invariance and thermodynamic linearity requires an exhaustive characterization of the admissible functional space. The manuscript must explicitly derive why alternatives (e.g., other linear combinations or scale-invariant penalties preserving thermodynamic linearity) are excluded, rather than relying on empirical validation alone; without this, the uniqueness step is not demonstrated and the attractor claim in Theorem 3 does not follow as a mathematical necessity.
Authors: We agree that the current presentation of Theorem 2 relies in part on empirical validation for excluding alternatives and would benefit from a more exhaustive mathematical characterization. In the revision, we will derive the general form of admissible functionals by imposing scale invariance under the allometric rescaling group and thermodynamic linearity (additivity under independent subsystems). We will explicitly show that any functional satisfying these must reduce to a multiple of the fractional metabolic excess; other candidates, such as logarithmic or alternative linear combinations, either break linearity or introduce scale-dependent terms that violate the gauge invariance. This will place uniqueness on a purely mathematical basis. revision: yes
-
Referee: [Theorem 3] Theorem 3: The claim that η* is the unique minimax attractor for the allometric class A(G,p,α_w) rests on the uniqueness established in Theorem 2. If other admissible functionals exist that remain dimensionless and linear, the incommensurability argument no longer forces a single attractor, undermining the universality of α* ≈ 2.72 as a necessity rather than a possible outcome.
Authors: Once the uniqueness of the cost functional is established mathematically in the revised Theorem 2, the minimax attractor property follows as a direct consequence of the incommensurability. The architectural invariance theorem already demonstrates that η* is the sole quantity invariant under the allometric class transformations that can simultaneously extremize both the now-unique metabolic gauge and the wave-reflection penalties. We will add an explicit corollary in the revision linking the two theorems to clarify that no other attractor is possible within the admissible class. revision: partial
-
Referee: [Theorem 1] No-Go Theorem (Theorem 1): The demonstration that local optimization requires a coupling parameter varying by 10^2–10^3 across the hierarchy must be shown to be independent of the specific choice of cost functional; otherwise the no-go result may be an artifact of the particular penalties chosen rather than a general consequence of incommensurability.
Authors: The No-Go Theorem is derived from the fundamental scaling mismatch between any extensive cost (which acquires a dimensional factor under hierarchical rescaling) and any dimensionless penalty. This dimensional incommensurability necessitates a hierarchy-dependent coupling regardless of the precise functional details, provided the costs retain their extensive versus dimensionless character. To make this generality explicit, the revision will include a short general argument using only dimensional analysis and the gauge-invariance properties, showing that the 10^2–10^3 variation in the coupling parameter is unavoidable for any such pair of costs. revision: yes
Circularity Check
No significant circularity in the derivation chain
full rationale
The paper structures its argument as three sequential theorems deriving from stated physical constraints (scale invariance, thermodynamic linearity, incommensurable costs). Theorem 2 asserts uniqueness of the fractional metabolic excess functional, and Theorem 3 derives the minimax invariant from the resulting allometric class. No equation in the provided abstract or description reduces by construction to a prior fitted input or self-referential definition; the claims are presented as proofs rather than renamings or tautologies. The derivation remains self-contained against the external benchmarks of allometric data and prior single-mechanism models.
Axiom & Free-Parameter Ledger
free parameters (1)
- coupling parameter
axioms (2)
- domain assumption Thermodynamic linearity and scale invariance uniquely determine the dimensionless cost functional as the fractional metabolic excess.
- domain assumption The allometric class A(G,p,α_w) is independent of absolute metabolic scales.
Reference graph
Works this paper leans on
-
[1]
G. S. Kassab, C. A. Rider, N. J. Tang, and Y. C. Fung. Morphometry of pig coronary arterial trees.Am. J. Physiol., 265:H350–H365, 1993. https://doi.org/10.1152/ajpheart.1993. 265.1.H350
-
[2]
M. Zamir. On fractal properties of arterial trees.J. Theor. Biol., 197:517–526, 1999. https: //doi.org/10.1006/jtbi.1998.0892. 9
- [3]
- [4]
-
[5]
The Dynamic Origin of Kleiber's Law
R. Marchesi. The dynamic origin of Kleiber’s law and the generalized metabolic scaling theorem.arXiv preprint, 2026.https://arxiv.org/abs/2604.10476
work page internal anchor Pith review Pith/arXiv arXiv 2026
-
[6]
C. D. Murray. The physiological principle of minimum work.Proc. Natl. Acad. Sci. USA, 12:207–214, 1926.https://doi.org/10.1073/pnas.12.3.207
-
[7]
T. F. Sherman. On connecting large vessels to small: the meaning of Murray’s law.J. Gen. Physiol., 78:431–453, 1981.https://doi.org/10.1085/jgp.78.4.431
-
[8]
G. B. West, J. H. Brown, and B. J. Enquist. A general model for the origin of allometric scaling laws in biology.Science, 276:122–126, 1997. https://doi.org/10.1126/science. 276.5309.122
-
[9]
J. Bennett. Murray’s law as an entropy-per-information-cost extremum.arXiv preprint, 2025. https://doi.org/10.48550/arXiv.2511.04022
-
[10]
J. Bennett. A single-index theory of optimal branching.arXiv preprint, 2025. https: //doi.org/10.48550/arXiv.2511.19915
-
[11]
L. Onsager. Reciprocal relations in irreversible processes.Phys. Rev., 37:405–426, 1931. https://doi.org/10.1103/PhysRev.37.405
-
[12]
Prince- ton University Press, Princeton, NJ, 2009
Aharon Ben-Tal, Laurent El Ghaoui, and Arkadi Nemirovski.Robust Optimization. Princeton University Press, 2009.https://doi.org/10.1515/9781400831050
-
[13]
Mathematische Annalen , year =
J. von Neumann. Zur Theorie der Gesellschaftsspiele.Math. Ann., 100:295–320, 1928. https://doi.org/10.1007/BF01448847
-
[14]
A. Forjaz, A. L. Kiemen, and D. Wirtz. Whole organism 3D mapping reveals universal branching topology and biophysical optimization governs vascular and nervous system development.bioRxiv preprint, 2026.https://doi.org/10.64898/2026.04.10.717729
-
[15]
Prigogine.Introduction to Thermodynamics of Irreversible Processes
I. Prigogine.Introduction to Thermodynamics of Irreversible Processes. Wiley, New York, 3rd edition, 1967. 10
1967
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.