Recognition: 2 theorem links
· Lean TheoremCellular-scale mechanism of cell crawling responding to substrate stiffness
Pith reviewed 2026-05-11 00:50 UTC · model grok-4.3
The pith
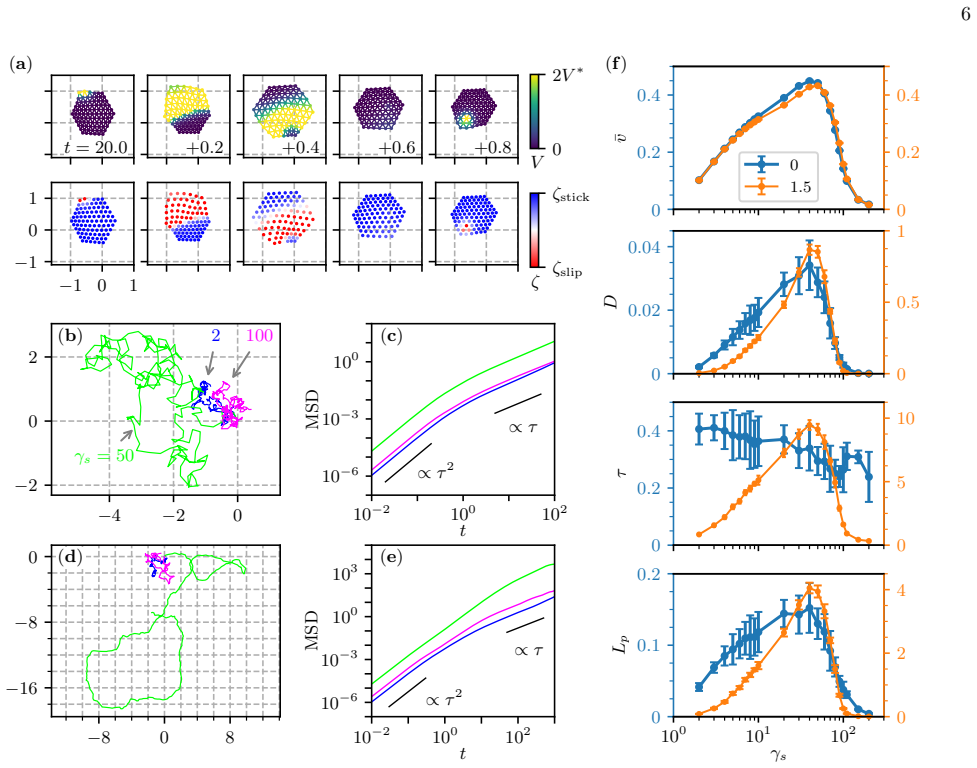
Cells migrate fastest and most persistently on substrates of intermediate stiffness according to a mechanochemical model of crawling.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
A mechanochemical model that integrates intracellular biochemical reactions with cellular deformation and substrate adhesion predicts that characteristic speed and diffusion constant of crawling cells vary non-monotonically with substrate stiffness, producing an optimal stiffness for migration; addition of a memory effect that feeds cell mechanics back to the chemical reactions further makes persistence time increase with stiffness on substrates softer than the optimum.
What carries the argument
Mechanochemical model coupling intracellular biochemical reactions to cell deformation and substrate adhesion, with an optional memory feedback term from mechanics to chemistry.
If this is right
- Cells should exhibit a stiffness optimum for efficient migration even in the absence of chemical gradients.
- Disrupting the mechanical-to-chemical feedback loop should eliminate the increase in persistence on soft substrates.
- Cell shape asymmetry and adhesion site distribution should show a non-monotonic dependence on stiffness that matches the motility curves.
- The same framework can be used to explore how changing adhesion strength or reaction rates shifts the location of the optimal stiffness.
Where Pith is reading between the lines
- The non-monotonic motility curve could explain why some cell types prefer particular tissue stiffnesses in wound healing or tumour invasion.
- Testing the model on cells with altered myosin activity or integrin expression would isolate which mechanical parameters control the position of the optimum.
- The memory effect suggests that cells retain a short-term record of recent substrate stiffness, which could be probed by suddenly changing stiffness during migration.
Load-bearing premise
The model assumes that mechanical feedback from cell deformation to intracellular chemistry can be represented by a simple memory effect and that this feedback alone accounts for the observed rise in persistence on soft substrates.
What would settle it
Measure the mean speed, diffusion constant, and persistence time of cells on a series of substrates whose stiffnesses bracket the predicted optimum and check whether speed and diffusion peak together while persistence rises only on the softer side.
Figures



read the original abstract
Biological cells are able to adapt their behaviour in response to environmental cues. Durotaxis is a phenomenon in which cells adjust their migration depending on the mechanical properties of a surrounding substrate. Although durotaxis has been studied more than two decades, basic cellular-scale mechanism of how cells regulate the motility responding to substrate stiffness remains to be elucidated. We address this issue by developing a theory utilising a mechanochemical model that integrates intracellular biochemical reactions with cellular deformation and substrate adhesion. Numerical analysis reveals that the characteristic speed and diffusion constant of cells change non-monotonically with respect to substrate stiffness, indicating the emergence of an optimal stiffness for migration. In addition, by introducing a memory effect that allows feedback from cell mechanics to the intracellular chemical reactions, the persistence time increases with substrate stiffness on a substrate softer than the optimal. We further investigate theoretically the origin of the non-monotonic dependence, that is comparable to the experimental observations, in terms of cell deformation and symmetry breaking in substrate adhesion. We believe that our study provides a unifying framework to understand complex durotactic cell migration.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript develops a mechanochemical model coupling intracellular biochemical reactions, cellular deformation, and substrate adhesion to investigate durotaxis. Numerical analysis demonstrates non-monotonic dependence of cell speed and diffusion constant on substrate stiffness, identifying an optimal stiffness for migration. A memory effect enabling feedback from mechanics to chemistry is shown to increase persistence time on substrates softer than optimal. The non-monotonicity is analyzed theoretically via cell deformation and symmetry breaking in adhesion, with results stated to be comparable to experiments.
Significance. If the numerical results hold, the work supplies a unifying cellular-scale framework linking mechanochemical feedback to durotactic behaviors, including optimal stiffness and stiffness-dependent persistence. The explicit treatment of deformation and adhesion symmetry breaking offers a mechanistic explanation that aligns with known experimental trends in cell motility on stiffness gradients.
minor comments (3)
- The abstract states that results are 'comparable to the experimental observations' but provides no citations or quantitative metrics; the main text should include specific experimental references and, if possible, direct overlays or tables comparing model outputs to data.
- The implementation of the memory effect (feedback from mechanics to intracellular reactions) is central to the persistence-time claim; its mathematical form, parameter values, and numerical integration scheme should be presented with sufficient detail for reproducibility.
- Figure captions and axis labels for the non-monotonic curves should explicitly state the range of stiffness values, the definition of 'characteristic speed' and 'diffusion constant,' and any averaging procedures used.
Simulated Author's Rebuttal
We thank the referee for their positive summary of our manuscript, recognition of its significance in providing a unifying mechanochemical framework for durotaxis, and recommendation for minor revision. No specific major comments were listed in the report, so we have no individual points requiring point-by-point rebuttal or revision at this stage. We remain available to address any additional minor issues the editor or referee may identify.
Circularity Check
No significant circularity; numerical results emerge from explicit model dynamics
full rationale
The paper constructs a mechanochemical model coupling intracellular reactions, cell deformation, and adhesion, then solves it numerically to obtain non-monotonic speed/diffusion versus stiffness. The memory effect is introduced explicitly as an additional term rather than derived from prior self-citation or fitted inputs. No equations reduce by construction to their own outputs, no parameters are tuned on the target observables and then relabeled as predictions, and no uniqueness theorem or ansatz is smuggled via self-citation. The derivation chain is therefore self-contained against external benchmarks.
Axiom & Free-Parameter Ledger
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclearNumerical analysis reveals that the characteristic speed and diffusion constant of cells change non-monotonically with respect to substrate stiffness... by introducing a memory effect that allows feedback from cell mechanics to the intracellular chemical reactions
-
IndisputableMonolith/Foundation/RealityFromDistinction.leanreality_from_one_distinction unclearsubcellular-element model... visco-elastic springs of Kelvin-Voigt type... Gray-Scott type reaction-diffusion equations
Reference graph
Works this paper leans on
-
[1]
Nakagaki, H
T. Nakagaki, H. Yamada, and ´A. T´ oth, Maze-solving by an amoeboid organism, Nature407, 470 (2000)
2000
-
[2]
J. S. King and R. H. Insall, Chemotaxis: finding the way forward with dictyostelium, Trends in cell biology19, 523 10 (2009)
2009
-
[3]
Hegemann, W
P. Hegemann, W. G¨ artner, and R. Uhl, All-trans reti- nal constitutes the functional chromophore in chlamy- domonas rhodopsin, Biophysical journal60, 1477 (1991)
1991
-
[4]
K. L. Poff and M. Skokut, Thermotaxis by pseudoplas- modia of dictyostelium discoideum., Proceedings of the National Academy of Sciences74, 2007 (1977)
2007
-
[5]
H¨ ader and R
D.-P. H¨ ader and R. Hemmersbach, Gravitaxis in euglena, Euglena: biochemistry, cell and molecular biology , 237 (2017)
2017
-
[6]
E. F. DeLong, R. B. Frankel, and D. A. Bazylinski, Multi- ple evolutionary origins of magnetotaxis in bacteria, Sci- ence259, 803 (1993)
1993
-
[7]
E. M. Purcell, Life at low Reynolds number, American Journal of Physics45, 3 (1977)
1977
-
[8]
Tanimoto and M
H. Tanimoto and M. Sano, A simple force-motion rela- tion for migrating cells revealed by multipole analysis of traction stress, Biophysical journal106, 16 (2014)
2014
-
[9]
Danuser, J
G. Danuser, J. Allard, and A. Mogilner, Mathematical modeling of eukaryotic cell migration: Insights beyond experiments, Annual Review of Cell and Developmental Biology29, 501 (2013)
2013
-
[10]
Leoni and P
M. Leoni and P. Sens, Model of cell crawling controlled by mechanosensitive adhesion, Phys. Rev. Lett.118, 228101 (2017)
2017
-
[11]
Tarama and R
M. Tarama and R. Yamamoto, Mechanics of cell crawl- ing by means of force-free cyclic motion, journal of the physical society of japan87, 044803 (2018)
2018
-
[12]
Lo, H.-B
C.-M. Lo, H.-B. Wang, M. Dembo, and Y.-l. Wang, Cell movement is guided by the rigidity of the substrate, Bio- physical journal79, 144 (2000)
2000
-
[13]
B. C. Isenberg, P. A. DiMilla, M. Walker, S. Kim, and J. Y. Wong, Vascular smooth muscle cell durotaxis de- pends on substrate stiffness gradient strength, Biophysi- cal journal97, 1313 (2009)
2009
-
[14]
J. R. Tse and A. J. Engler, Stiffness gradients mimicking in vivo tissue variation regulate mesenchymal stem cell fate, PloS one6, e15978 (2011)
2011
-
[15]
E. B. Evans, S. W. Brady, A. Tripathi, and D. Hoffman- Kim, Schwann cell durotaxis can be guided by physiolog- ically relevant stiffness gradients, Biomaterials research 22, 14 (2018)
2018
-
[16]
B. J. DuChez, A. D. Doyle, E. K. Dimitriadis, and K. M. Yamada, Durotaxis by human cancer cells, Biophysical journal116, 670 (2019)
2019
-
[17]
C. Kang, P. Chen, X. Yi, D. Li, Y. Hu, Y. Yang, H. Cai, B. Li, and C. Wu, Amoeboid cells undergo durotaxis with soft end polarized nmiia, Elife13, RP96821 (2024)
2024
-
[18]
Isomursu, K.-Y
A. Isomursu, K.-Y. Park, J. Hou, B. Cheng, M. Mathieu, G. A. Shamsan, B. Fuller, J. Kasim, M. M. Mahmoodi, T. J. Lu,et al., Directed cell migration towards softer environments, Nature materials21, 1081 (2022)
2022
-
[19]
Huang, J
Y. Huang, J. Su, J. Liu, X. Yi, F. Zhou, J. Zhang, J. Wang, X. Meng, L. Si, and C. Wu, Yap activation in promoting negative durotaxis and acral melanoma pro- gression, Cells11, 3543 (2022)
2022
-
[20]
S. P. Singh, M. P. Schwartz, J. Y. Lee, B. D. Fairbanks, and K. S. Anseth, A peptide functionalized poly (ethy- lene glycol)(peg) hydrogel for investigating the influence of biochemical and biophysical matrix properties on tu- mor cell migration, Biomaterials science2, 1024 (2014)
2014
-
[21]
Ebata, K
H. Ebata, K. Moriyama, T. Kuboki, and S. Kidoaki, Gen- eral cellular durotaxis induced with cell-scale heterogene- ity of matrix-elasticity, Biomaterials230, 119647 (2020)
2020
-
[22]
Ebata and S
H. Ebata and S. Kidoaki, Interplay among cell migration, shaping, and traction force on a matrix with cell-scale stiffness heterogeneity, Biophysics and physicobiology19, e190036 (2022)
2022
-
[23]
Iwadate, C
Y. Iwadate, C. Okimura, K. Sato, Y. Nakashima, M. Tsu- jioka, and K. Minami, Myosin-ii-mediated directional mi- gration of dictyostelium cells in response to cyclic stretch- ing of substratum, Biophysical journal104, 748 (2013)
2013
-
[24]
Shellard and R
A. Shellard and R. Mayor, Collective durotaxis along a self-generated stiffness gradient in vivo, Nature600, 690 (2021)
2021
-
[25]
P. D. P. Dingal, A. M. Bradshaw, S. Cho, M. Raab, A. Buxboim, J. Swift, and D. E. Discher, Fractal het- erogeneity in minimal matrix models of scars modu- lates stiff-niche stem-cell responses via nuclear exit of a mechanorepressor, Nature materials14, 951 (2015)
2015
-
[26]
W. H. Goldmann, Durotaxis: A cause of organ fibrosis and metastatic cancer?, Cell Biology International48, 553 (2024)
2024
-
[27]
A. J. Engler, S. Sen, H. L. Sweeney, and D. E. Discher, Matrix elasticity directs stem cell lineage specification, cell126, 677 (2006)
2006
-
[28]
Shellard and R
A. Shellard and R. Mayor, Durotaxis: The hard path from in vitro to in vivo, Developmental Cell56, 227 (2021)
2021
-
[29]
B. L. Bangasser, S. S. Rosenfeld, and D. J. Odde, Deter- minants of maximal force transmission in a motor-clutch model of cell traction in a compliant microenvironment, Biophysical journal105, 581 (2013)
2013
-
[30]
M. E. Pallar` es, I. Pi-Jaum` a, I. C. Fortunato, V. Grazu, M. G´ omez-Gonz´ alez, P. Roca-Cusachs, J. M. de la Fuente, R. Alert, R. Sunyer, J. Casademunt,et al., Stiffness-dependent active wetting enables optimal col- lective cell durotaxis, Nature Physics19, 279 (2023)
2023
-
[31]
E. A. Novikova, M. Raab, D. E. Discher, and C. Storm, Persistence-driven durotaxis: Generic, directed motil- ity in rigidity gradients, Phys. Rev. Lett.118, 078103 (2017)
2017
-
[32]
T. J. Newman, Modeling multicellular structures using the subcellular element model, Single-cell-based models in biology and medicine , 221 (2007)
2007
-
[33]
Tarama, K
M. Tarama, K. Mori, and R. Yamamoto, Mechanochemi- cal subcellular-element model of crawling cells, Frontiers in Cell and Developmental Biology10, 1046053 (2022)
2022
-
[34]
Taniguchi, S
D. Taniguchi, S. Ishihara, T. Oonuki, M. Honda- Kitahara, K. Kaneko, and S. Sawai, Phase geometries of two-dimensional excitable waves govern self-organized morphodynamics of amoeboid cells, Proceedings of the National Academy of Sciences110, 5016 (2013)
2013
-
[35]
Koshizuka, A
S. Koshizuka, A. Nobe, and Y. Oka, Numerical analysis of breaking waves using the moving particle semi-implicit method, International journal for numerical methods in fluids26, 751 (1998)
1998
-
[36]
U. S. Schwarz and S. A. Safran, Physics of adherent cells, Reviews of Modern Physics85, 1327 (2013)
2013
-
[37]
Moriyama and S
K. Moriyama and S. Kidoaki, Cellular durotaxis re- visited: initial-position-dependent determination of the threshold stiffness gradient to induce durotaxis, Lang- muir35, 7478 (2018)
2018
-
[38]
J. A. Espina, C. L. Marchant, and E. H. Barriga, Duro- taxis: the mechanical control of directed cell migration, The FEBS journal289, 2736 (2022)
2022
-
[39]
J. R. Taylor,An introduction to error analysis: the study of uncertainties in physical measurements(MIT Press, 11 2022)
2022
-
[40]
S. P. Palecek, J. C. Loftus, M. H. Ginsberg, D. A. Lauffenburger, and A. F. Horwitz, Integrin-ligand bind- ing properties govern cell migration speed through cell- substratum adhesiveness, Nature385, 537 (1997)
1997
-
[41]
Schreiber, B
C. Schreiber, B. Amiri, J. C. J. Heyn, J. O. R adler, and M. Falcke, On the adhesion–velocity relation and length adaptation of motile cells on stepped fibronectin lanes, Proceedings of the National Academy of Sciences118, e2009959118 (2021)
2021
-
[42]
Missirlis and J
D. Missirlis and J. P. Spatz, Combined effects of peg hy- drogel elasticity and cell-adhesive coating on fibroblast adhesion and persistent migration, Biomacromolecules 15, 195 (2014)
2014
-
[43]
House, M
D. House, M. L. Walker, Z. Wu, J. Y. Wong, and M. Betke, Tracking of cell populations to understand their spatio-temporal behavior in response to physical stimuli, in2009 IEEE Computer Society Conference on Computer Vision and Pattern Recognition Workshops (IEEE, 2009) pp. 186–193
2009
-
[44]
P. W. Oakes, D. C. Patel, N. A. Morin, D. P. Zitter- bart, B. Fabry, J. S. Reichner, and J. X. Tang, Neutrophil morphology and migration are affected by substrate elas- ticity, Blood, The Journal of the American Society of Hematology114, 1387 (2009)
2009
-
[45]
Ji and Y
C. Ji and Y. Huang, Durotaxis and negative durotaxis: where should cells go?, Communications Biology6, 1169 (2023)
2023
-
[46]
Arellano-Tint´ o, D
J. Arellano-Tint´ o, D. Stepanova, H. M. Byrne, P. K. Maini, and T. Alarc´ on, Multiscale modelling shows how cell-ecm interactions impact ecm fibre alignment and cell detachment, PLOS Computational Biology21, 1 (2025)
2025
-
[47]
J. J. Molina and R. Yamamoto, Modeling the mechanosensitivity of fast-crawling cells on cyclically stretched substrates, Soft Matter15, 683 (2019)
2019
-
[48]
L¨ ober, F
J. L¨ ober, F. Ziebert, and I. S. Aranson, Modeling crawl- ing cell movement on soft engineered substrates, Soft Matter10, 1365 (2014)
2014
-
[49]
Fukushima, S
S. Fukushima, S. Matsuoka, and M. Ueda, Excitable dy- namics of ras triggers spontaneous symmetry breaking of pip3 signaling in motile cells, Journal of Cell Science 132, jcs224121 (2019)
2019
-
[50]
L. Li, S. F. Nørrelykke, and E. C. Cox, Persistent cell mo- tion in the absence of external signals: a search strategy for eukaryotic cells, PLoS one3, e2093 (2008)
2008
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.