Recognition: no theorem link
A Brownian dynamics study of liquid-liquid phase separation in multi-scale chromatin networks
Pith reviewed 2026-05-15 01:37 UTC · model grok-4.3
The pith
Chromatin fiber networks control the positioning and morphology of phase-separated protein droplets through interactions analogous to wetting transitions.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
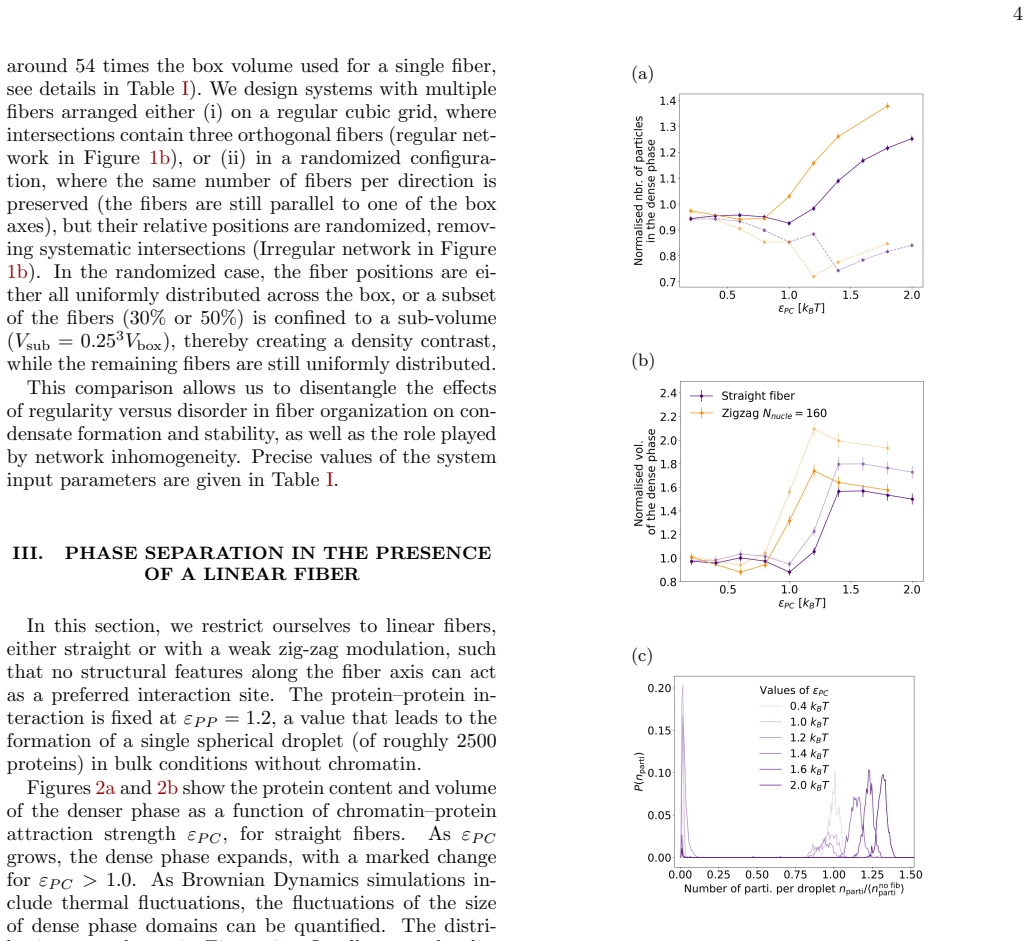
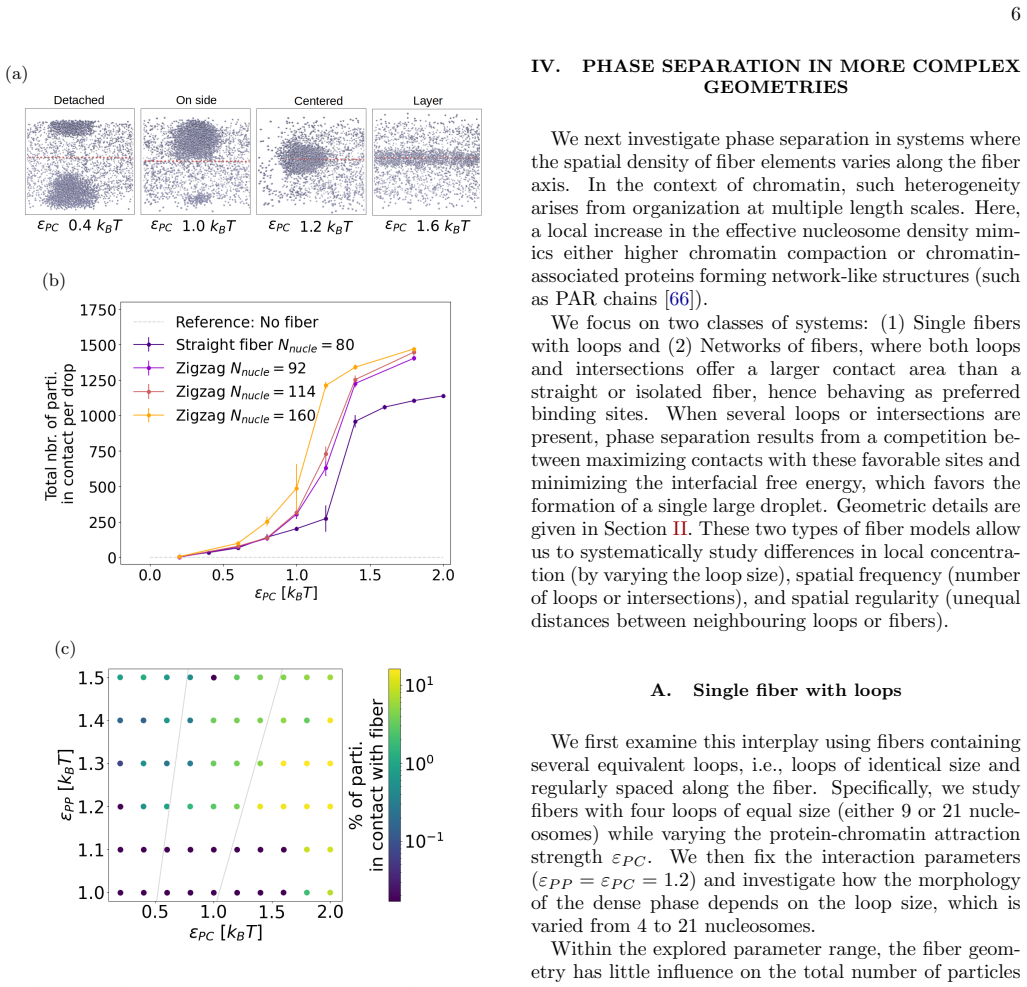
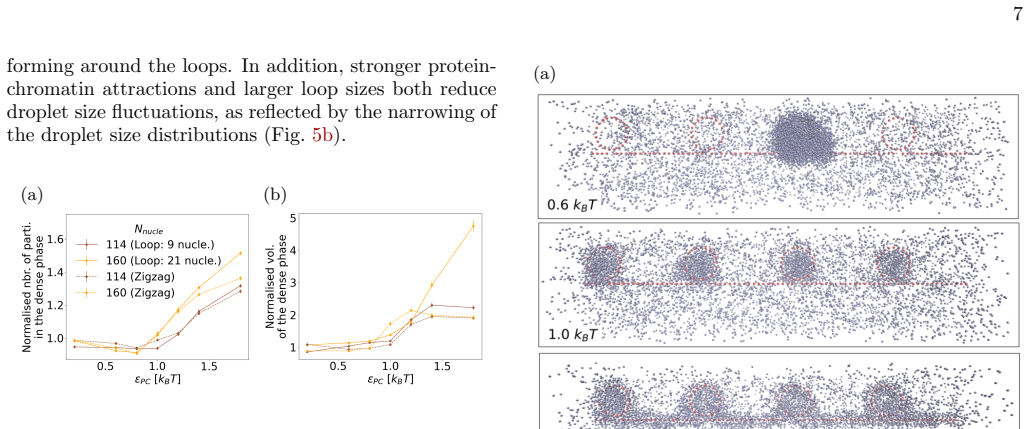
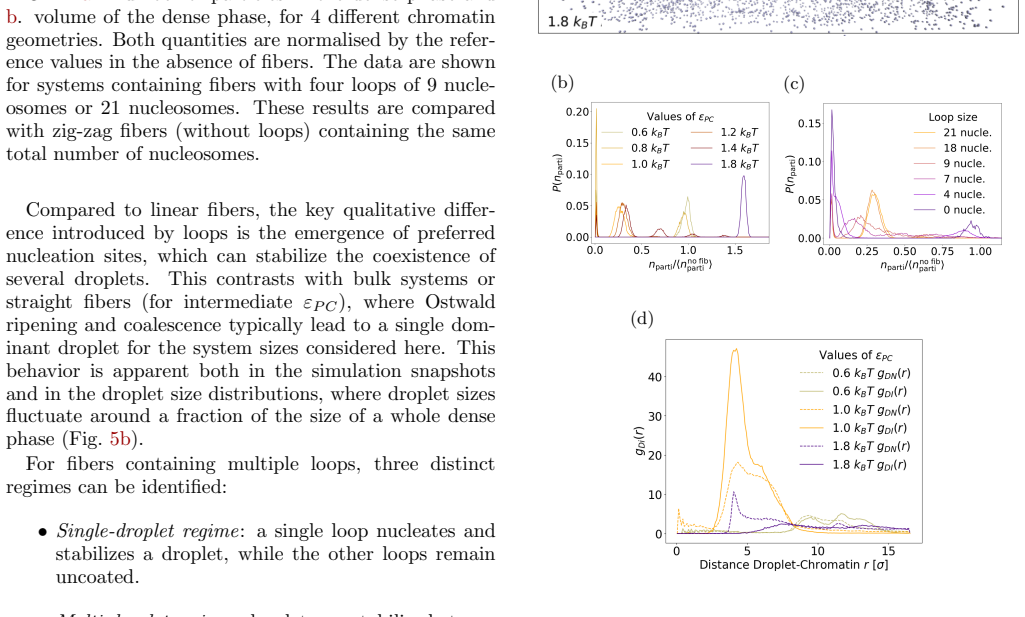
Using Brownian dynamics of Lennard-Jones particles on fixed fibrous substrates that represent multi-scale chromatin, the work shows that protein-fiber interactions determine droplet positioning in a manner directly analogous to wetting transitions. Both local geometric constraints from individual fibers and global network organization control droplet size, morphology, and multiplicity. Large-scale asymmetries in fiber arrangement further induce robust spatial localization of the dense phase.
What carries the argument
Minimal model of Lennard-Jones particles interacting with fixed fibrous substrates, which encodes the multiscale chromatin architecture and its effect on phase separation.
If this is right
- Droplet placement follows rules similar to wetting transitions controlled by fiber-protein attraction strength.
- Local fiber spacing and curvature set upper limits on droplet size and enforce specific shapes.
- Global network topology modulates the number of coexisting droplets.
- Asymmetric fiber distributions produce stable, non-uniform localization of the dense phase.
Where Pith is reading between the lines
- Altering fiber asymmetry through loop extrusion or other remodeling processes could reposition condensates without changing protein concentrations.
- The same physical rules may apply to other nuclear scaffolds such as RNA or PAR chains if they present comparable fibrous geometry.
- Chromatin compaction changes during the cell cycle could switch condensate multiplicity or localization as a direct physical consequence.
Load-bearing premise
A model with immobile fibers and simple particle interactions is enough to capture the dominant physical effects that real chromatin exerts on phase-separating proteins.
What would settle it
Direct imaging of droplet positions in cells or in vitro chromatin networks that either matches or deviates from the predicted wetting-like localization when fiber geometry or asymmetry is systematically varied.
Figures




read the original abstract
In living cells, proteins involved in specialized biochemical functions are often spatially organized within biomolecular condensates. Increasing evidence suggests that some of these condensates, including DNA repair condensates, emerge through liquid-liquid phase separation (LLPS). In the nucleus, however, condensates form within a highly heterogeneous environment composed of chromatin fibers, RNA, and additional protein scaffolds such as PAR chains, all of which may interact with phase-separating proteins. Moreover, condensate formation is frequently associated with specific chromatin conformations; for instance, loop extrusion has been proposed as a mechanism promoting DNA repair condensates. Here, we investigate how the surrounding fibrous environment controls the morphology and spatial organization of phase-separated condensates. Using Brownian dynamics simulations of minimal models combining Lennard-Jones particles with fixed fibrous substrates, we examine the respective roles of local fiber geometry and large-scale network organization, reflecting the multiscale architecture of chromatin. We show that protein-fiber interactions strongly influence droplet positioning relative to the substrate, in a manner analogous to wetting transitions in soft condensed matter systems. Both local geometric constraints and global network organization markedly affect droplet size, morphology, and multiplicity. In addition, large-scale asymmetries in fiber organization can induce robust spatial localization of the dense phase. Our results thus highlight how multiscale structural heterogeneity of the nuclear environment can regulate the emergence and organization of biomolecular condensates.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
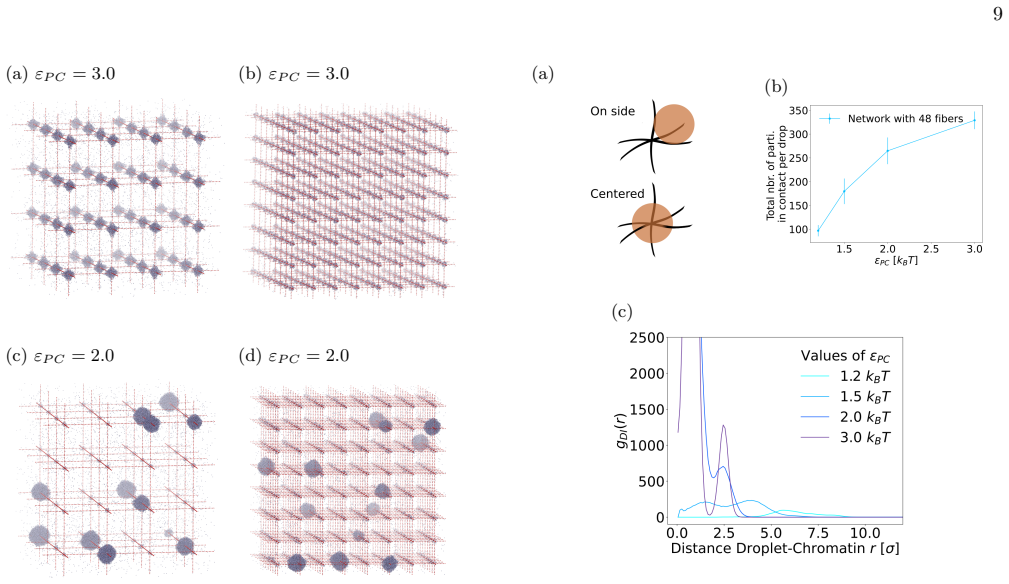
Summary. The paper presents Brownian dynamics simulations of Lennard-Jones particles on fixed fibrous substrates modeling multi-scale chromatin networks. It claims that protein-fiber interaction strength controls droplet positioning in a manner analogous to wetting transitions, that local geometric constraints and global network organization control droplet size, morphology and multiplicity, and that large-scale fiber asymmetries induce robust spatial localization of the dense phase.
Significance. If the minimal fixed-fiber model captures the dominant physics, the results provide a clear computational demonstration that nuclear architectural heterogeneity can regulate condensate positioning and morphology via mechanisms familiar from soft-matter wetting. The direct simulation approach with only two free parameters (interaction strength and fiber density/connectivity) is a strength, as are the explicit comparisons of local versus global network effects.
major comments (3)
- [Methods] Methods (simulation setup): all results, including the central claim of robust localization induced by large-scale asymmetries, are obtained with fibers held completely fixed. The manuscript contains no runs or discussion of mobile fibers that could respond to condensate-induced stresses; this omission is load-bearing for the extrapolation to real chromatin.
- [Results] Results (wetting and positioning sections): the analogy to wetting transitions is stated qualitatively but no quantitative benchmarks are given (e.g., measured contact angles, critical interaction strengths, or comparison to known wetting criteria on structured substrates).
- [Results] Results (asymmetry and localization figures): reported trends in droplet size, multiplicity, and spatial localization are presented without error bars, ensemble statistics, or finite-size scaling checks, making it impossible to assess the robustness asserted in the abstract and conclusions.
minor comments (2)
- [Abstract] Abstract: the phrase 'multi-scale chromatin networks' is used while the model employs static fibers; a brief clarification of which scales are represented would improve readability.
- [Figures] Figure captions: several panels lack explicit labels for the protein-fiber interaction parameter values used; adding these would aid reproducibility.
Simulated Author's Rebuttal
We thank the referee for their constructive and detailed comments, which have helped us identify areas for improvement in clarity and rigor. We address each major comment below and will revise the manuscript to incorporate the suggested enhancements while preserving the core findings of our minimal fixed-fiber model.
read point-by-point responses
-
Referee: [Methods] Methods (simulation setup): all results, including the central claim of robust localization induced by large-scale asymmetries, are obtained with fibers held completely fixed. The manuscript contains no runs or discussion of mobile fibers that could respond to condensate-induced stresses; this omission is load-bearing for the extrapolation to real chromatin.
Authors: We agree that the fibers are held fixed in our simulations, which is a deliberate modeling choice to isolate the effects of multi-scale geometry and network organization on condensate behavior. This minimal approach allows direct attribution of positioning, size, and morphology trends to fiber architecture without confounding dynamics. While real chromatin can exhibit mobility, the fixed-fiber limit is appropriate for exploring architectural control in a computationally tractable setting. In the revision we will add an explicit limitations subsection discussing the fixed-fiber approximation, its relation to semi-rigid chromatin, and the potential effects of fiber mobility as a direction for future work. revision: partial
-
Referee: [Results] Results (wetting and positioning sections): the analogy to wetting transitions is stated qualitatively but no quantitative benchmarks are given (e.g., measured contact angles, critical interaction strengths, or comparison to known wetting criteria on structured substrates).
Authors: We thank the referee for this observation. In the revised manuscript we will quantify the wetting analogy by extracting contact angles from the simulated droplet interfaces, determining the critical interaction strength at which the positioning transition occurs, and comparing these values to established soft-matter criteria for wetting on structured and heterogeneous substrates. These additions will be placed in the wetting and positioning results sections to make the analogy rigorous. revision: yes
-
Referee: [Results] Results (asymmetry and localization figures): reported trends in droplet size, multiplicity, and spatial localization are presented without error bars, ensemble statistics, or finite-size scaling checks, making it impossible to assess the robustness asserted in the abstract and conclusions.
Authors: We acknowledge that the current figures lack statistical detail. In the revision we will add error bars (standard deviations from at least five independent runs per condition) to all reported droplet properties and include ensemble averages over multiple network realizations. We will also perform limited system-size checks on larger networks to support the robustness claims. A comprehensive finite-size scaling study is computationally intensive and will be noted as future work, but the added statistics will allow readers to evaluate the trends directly. revision: partial
Circularity Check
No circularity: direct numerical outcomes from explicit minimal model
full rationale
The paper reports results exclusively from Brownian dynamics simulations of Lennard-Jones particles interacting with fixed fibrous substrates. All observations on droplet positioning, size, morphology, multiplicity, and asymmetry-driven localization are obtained by direct integration of the stochastic equations of motion under the chosen potentials; no quantities are obtained by fitting parameters to a subset of data and then relabeling them as predictions, no equations are defined in terms of their own outputs, and no load-bearing claims rest on self-citations that themselves reduce to ansatz or prior fitted results. The fixed-fiber choice is stated explicitly as a modeling simplification whose effects are explored numerically rather than derived tautologically. The derivation chain is therefore self-contained against external benchmarks and contains no reductions of the enumerated kinds.
Axiom & Free-Parameter Ledger
free parameters (2)
- protein-fiber interaction strength
- fiber density and connectivity parameters
axioms (2)
- standard math Overdamped Brownian dynamics governs particle motion on the relevant timescales
- domain assumption Fibers remain completely fixed and do not respond to droplet forces
Reference graph
Works this paper leans on
-
[1]
Linear fiber There is a large variety of chromatin geometries within the length scales under study here (nucleosomes to con- densates, 10 nm to 1µm). In order to understand the possible interactions of chromatin with condensate- forming proteins, we choose to concentrate on generic variations of the surface seen by condensate proteins near chromatin. We u...
-
[2]
There can be intersections within the same fiber if there is a loop [59, 60], or between dif- ferent fibers
Fibers with loops and networks We then then progressively increase the geometrical complexity by adding two related elements: (1) inter- sections, defined as regions of space where two nucleo- somes, which are not neighbors in the chain, come close to each other, and (2) the presence of several fibers in the simulation box. There can be intersections with...
-
[3]
Droplet identification Two proteins are considered as part of the same droplet if they are closer than a threshold distancer cut of each other. The value ofr cut is chosen as the distance at 12 the first minimum in the protein-protein pair correlation function (rcut = 1.65σfor the small systems with a single fiber andr cut = 1.55σfor the large systems wit...
-
[4]
It is estimated from the positionsr i = (rix, riy, riz) of the nparti proteins in each droplet
Estimating the droplet volume Quantifying the volume of the distinct phases within a microscopic system presents significant challenges due to the absence of clearly defined phase boundaries. It is estimated from the positionsr i = (rix, riy, riz) of the nparti proteins in each droplet. The center of mass of their positions along themcoordinate is calcula...
-
[5]
Droplet localization To quantify the localization of the droplets with re- spect to the fibers, we calculate the pair correlation func- tiong DN (r) between the center of mass of a droplet and the positions of all nucleosomes as: gDN (r) = * 1 NdropNnucle NdropX i NnucleX j δ(dij −r) + t ,(A4) dij being the distance between thei th droplet center of mass ...
-
[6]
C. P. Brangwynne, C. R. Eckmann, D. S. Courson, A. Rybarska, C. Hoege, J. Gharakhani, F. J¨ ulicher, and A. A. Hyman, Germline P Granules Are Liquid Droplets That Localize by Controlled Dissolution/Condensation, Science324, 1729 (2009)
2009
-
[7]
Y. Shin and C. P. Brangwynne, Liquid phase con- densation in cell physiology and disease, Science357, 10.1126/science.aaf4382 (2017)
-
[8]
Shin, Y.-C
Y. Shin, Y.-C. Chang, D. S. Lee, J. Berry, D. W. Sanders, P. Ronceray, N. S. Wingreen, M. Haataja, and C. P. Brangwynne, Liquid Nuclear Condensates Mechan- ically Sense and Restructure the Genome, Cell175, 1481 (2018)
2018
-
[9]
B. A. Gibson, L. K. Doolittle, M. W. Schneider, L. E. Jensen, N. Gamarra, L. Henry, D. W. Gerlich, S. Red- ding, and M. K. Rosen, Organization of Chromatin by Intrinsic and Regulated Phase Separation, Cell179, 470 (2019)
2019
-
[10]
C. A. Weber, D. Zwicker, F. J¨ ulicher, and C. F. Lee, Physics of active emulsions, Reports on Progress in Physics82, 064601 (2019)
2019
-
[11]
B. R. Sabari, A. Dall’Agnese, and R. A. Young, Biomolec- ular Condensates in the Nucleus, Trends in Biochemical Sciences45, 961 (2020)
2020
-
[12]
Gouveia, Y
B. Gouveia, Y. Kim, J. W. Shaevitz, S. Petry, H. A. Stone, and C. P. Brangwynne, Capillary forces generated by biomolecular condensates, Nature609, 255 (2022)
2022
-
[13]
M. N. Rouches and B. B. Machta, Polymer Collapse & Liquid-Liquid Phase-Separation are Coupled in a Gener- alized Prewetting Transition (2024)
2024
-
[14]
Hnisz, K
D. Hnisz, K. Shrinivas, R. A. Young, A. K. Chakraborty, and P. A. Sharp, A Phase Separation Model for Tran- scriptional Control, Cell169, 13 (2017)
2017
-
[15]
Rippe, Liquid–Liquid Phase Separation in Chromatin, Cold Spring Harbor Perspectives in Biology14, a040683 (2022)
K. Rippe, Liquid–Liquid Phase Separation in Chromatin, Cold Spring Harbor Perspectives in Biology14, a040683 (2022)
2022
-
[16]
J. Wu, B. Chen, Y. Liu, L. Ma, W. Huang, and Y. Lin, Modulating gene regulation function by chemically con- trolled transcription factor clustering, Nature Communi- cations13, 2663 (2022)
2022
-
[17]
Mann and D
R. Mann and D. Notani, Transcription factor condensates and signaling driven transcription, Nucleus14, 2205758 (2023)
2023
-
[18]
Brangwynne, P
C. Brangwynne, P. Tompa, and R. Pappu, Polymer physics of intracellular phase transitions, Nature Physics 11, 899 (2015)
2015
-
[19]
S. F. Banani, H. O. Lee, A. A. Hyman, and M. K. Rosen, Biomolecular condensates: organizers of cellular biochemistry, Nature Reviews Molecular Cell Biology18, 285 (2017)
2017
-
[20]
Kilic, A
S. Kilic, A. Lezaja, M. Gatti, E. Bianco, J. Michelena, R. Imhof, and M. Altmeyer, Phase separation of 53BP1 14 determines liquid-like behavior of DNA repair compart- ments, The EMBO Journal38, e101379 (2019)
2019
-
[21]
A. A. Hyman, C. A. Weber, and F. J¨ ulicher, Liquid- Liquid Phase Separation in Biology, Annual Review of Cell and Developmental Biology30, 39 (2014)
2014
-
[22]
Alberti, A
S. Alberti, A. Gladfelter, and T. Mittag, Considerations and Challenges in Studying Liquid-Liquid Phase Sep- aration and Biomolecular Condensates, Cell176, 419 (2019)
2019
-
[23]
M. L. Heltberg, J. Min´ e-Hattab, A. Taddei, A. M. Wal- czak, and T. Mora, Physical observables to determine the nature of membrane-less cellular sub-compartments, eLife10, e69181 (2021)
2021
-
[24]
Min´ e-Hattab, M
J. Min´ e-Hattab, M. Heltberg, M. Villemeur, C. Guedj, T. Mora, A. M. Walczak, M. Dahan, and A. Taddei, Sin- gle molecule microscopy reveals key physical features of repair foci in living cells, eLife10, e60577 (2021)
2021
-
[25]
M. S. Heltberg, A. Lucchetti, F.-S. Hsieh, D. P. Minh Nguyen, S.-h. Chen, and M. H. Jensen, Enhanced DNA repair through droplet formation and p53 oscilla- tions, Cell185, 4394 (2022)
2022
-
[26]
Garc´ ıa Fern´ andez, S
F. Garc´ ıa Fern´ andez, S. Huet, and J. Min´ e-Hattab, Multi-Scale Imaging of the Dynamic Organization of Chromatin, International Journal of Molecular Sciences 24, 15975 (2023)
2023
-
[27]
G. J. Filion, J. G. Van Bemmel, U. Braunschweig, W. Talhout, J. Kind, L. D. Ward, W. Brugman, I. J. De Castro, R. M. Kerkhoven, H. J. Bussemaker, and B. van Steensel, Systematic Protein Location Mapping Reveals Five Principal Chromatin Types in Drosophila Cells, Cell143, 212 (2010)
2010
-
[28]
A. N. Boettiger, B. Bintu, J. R. Moffitt, S. Wang, B. J. Beliveau, G. Fudenberg, M. Imakaev, L. A. Mirny, C.- t. Wu, and X. Zhuang, Super-resolution imaging reveals distinct chromatin folding for different epigenetic states, Nature529, 418 (2016)
2016
-
[29]
D. I. Cattoni, A. M. Cardozo Gizzi, M. Georgieva, M. Di Stefano, A. Valeri, D. Chamousset, C. Houbron, S. D´ ejardin, J.-B. Fiche, I. Gonz´ alez, J.-M. Chang, T. Sexton, M. A. Marti-Renom, F. Bantignies, G. Cav- alli, and M. Nollmann, Single-cell absolute contact prob- ability detection reveals chromosomes are organized by multiple low-frequency yet speci...
2017
-
[30]
Szabo, D
Q. Szabo, D. Jost, J.-M. Chang, D. I. Cattoni, G. L. Pa- padopoulos, B. Bonev, T. Sexton, J. Gurgo, C. Jacquier, M. Nollmann, F. Bantignies, and G. Cavalli, TADs are 3D structural units of higher-order chromosome organiza- tion inDrosophila, Science Advances4, eaar8082 (2018)
2018
-
[31]
Sexton, E
T. Sexton, E. Yaffe, E. Kenigsberg, F. Bantignies, B. Leblanc, M. Hoichman, H. Parrinello, A. Tanay, and G. Cavalli, Three-Dimensional Folding and Functional Organization Principles of the Drosophila Genome, Cell 148, 458 (2012)
2012
-
[32]
Arnould and G
C. Arnould and G. Legube, The Secret Life of Chromo- some Loops upon DNA Double-Strand Break, Journal of Molecular Biology432, 724 (2020)
2020
-
[33]
Arnould, V
C. Arnould, V. Rocher, A.-L. Finoux, T. Clouaire, K. Li, F. Zhou, P. Caron, P. E. Mangeot, E. P. Ricci, R. Mourad, J. E. Haber, D. Noordermeer, and G. Legube, Loop extrusion as a mechanism for formation of DNA damage repair foci, Nature590, 660 (2021)
2021
-
[34]
Arnould, V
C. Arnould, V. Rocher, F. Saur, A. S. Bader, F. Muz- zopappa, S. Collins, E. Lesage, B. Le Bozec, N. Puget, T. Clouaire, T. Mangeat, R. Mourad, N. Ahituv, D. Noordermeer, F. Erdel, M. Bushell, A. Marnef, and G. Legube, Chromatin compartmentalization regulates the response to DNA damage, Nature623, 183 (2023)
2023
-
[35]
Shin and A
J. Shin and A. B. Kolomeisky, Facilitation of DNA loop formation by protein-DNA non-specific interactions, Soft Matter15, 5255 (2019)
2019
-
[36]
F. C. Keber, T. Nguyen, A. Mariossi, C. P. Brang- wynne, and M. W¨ uhr, Evidence for widespread cytoplas- mic structuring into mesoscale condensates, Nature Cell Biology26, 346 (2024)
2024
-
[37]
C. P. Broedersz, X. Wang, Y. Meir, J. J. Loparo, D. Z. Rudner, and N. S. Wingreen, Condensation and local- ization of the partitioning protein ParB on the bacterial chromosome, Proceedings of the National Academy of Sciences111, 8809 (2014)
2014
-
[38]
Zwicker, The intertwined physics of active chemical reactions and phase separation, Current Opinion in Col- loid & Interface Science61, 101606 (2022)
D. Zwicker, The intertwined physics of active chemical reactions and phase separation, Current Opinion in Col- loid & Interface Science61, 101606 (2022)
2022
-
[39]
A. R. Strom, J. M. Eeftens, Y. Polyachenko, C. J. Weaver, H.-F. Watanabe, D. Bracha, N. D. Orlovsky, C. C. Jumper, W. M. Jacobs, and C. P. Brangwynne, In- terplay of condensation and chromatin binding underlies BRD4 targeting, Molecular Biology of the Cell35, ar88 (2024)
2024
-
[40]
M. Du, S. H. Stitzinger, J.-H. Spille, W.-K. Cho, C. Lee, M. Hijaz, A. Quintana, and I. I. Ciss´ e, Direct observation of a condensate effect on super-enhancer controlled gene bursting, Cell187, 331 (2024)
2024
-
[41]
Rayleigh, On The Instability Of Jets, Proceedings of the London Mathematical Societys1-10, 4 (1878)
L. Rayleigh, On The Instability Of Jets, Proceedings of the London Mathematical Societys1-10, 4 (1878)
-
[42]
De Gennes, F
P.-G. De Gennes, F. Brochard-Wyart, and D. Qu´ er´ e, Capillarity and Wetting Phenomena(Springer New York, New York, NY, 2004)
2004
-
[43]
Eggers and E
J. Eggers and E. Villermaux, Physics of liquid jets, Re- ports on Progress in Physics71, 036601 (2008)
2008
-
[44]
Zhang, J
Y. Zhang, J. Sprittles, and D. Lockerby, Thermal capil- lary wave growth and surface roughening of nanoscale liq- uid films, Journal of Fluid Mechanics915, A135 (2021)
2021
-
[45]
Gopan and S
N. Gopan and S. P. Sathian, Rayleigh instability at small length scales, Physical Review E90, 033001 (2014)
2014
-
[46]
Zhang, J
Y. Zhang, J. E. Sprittles, and D. A. Lockerby, Nanoscale thin-film flows with thermal fluctuations and slip, Phys- ical Review E102, 053105 (2020)
2020
-
[47]
P. J. Flory, Thermodynamics of High Polymer Solutions, The Journal of Chemical Physics10, 51 (1942)
1942
-
[48]
M. L. Huggins, Some Properties of Solutions of Long- chain Compounds., The Journal of Physical Chemistry 46, 151 (1942)
1942
-
[49]
Tiani, M
R. Tiani, M. Jardat, and V. Dahirel, Phase transitions in chromatin: Mesoscopic and mean-field approaches, The Journal (2025)
2025
-
[50]
Ronceray, S
P. Ronceray, S. Mao, A. Koˇ smrlj, and M. P. Haataja, Liquid demixing in elastic networks: Cavitation, perme- ation, or size selection?, Europhysics Letters137, 67001 (2022)
2022
-
[51]
K. A. Rosowski, T. Sai, E. Vidal-Henriquez, D. Zwicker, R. W. Style, and E. R. Dufresne, Elastic ripening and in- hibition of liquid–liquid phase separation, Nature Physics 16, 422 (2020)
2020
-
[52]
R. W. Style, T. Sai, N. Fanelli, M. Ijavi, K. Smith- Mannschott, Q. Xu, L. A. Wilen, and E. R. Dufresne, Liquid-Liquid Phase Separation in an Elastic Network, Physical Review X8, 011028 (2018). 15
2018
-
[53]
D. S. W. Lee, N. S. Wingreen, and C. P. Brangwynne, Chromatin mechanics dictates subdiffusion and coarsen- ing dynamics of embedded condensates, Nature Physics 17, 531 (2021)
2021
-
[54]
Qi and B
Y. Qi and B. Zhang, Chromatin network retards nucleoli coalescence, Nature Communications12, 6824 (2021)
2021
-
[55]
M. P. Allen and D. J. Tildesley,Computer simulation of liquids, second edition ed. (Oxford University Press, Oxford, 2017)
2017
-
[56]
A. P. Thompson, H. M. Aktulga, R. Berger, D. S. Bolin- tineanu, W. M. Brown, P. S. Crozier, P. J. In ’T Veld, A. Kohlmeyer, S. G. Moore, T. D. Nguyen, R. Shan, M. J. Stevens, J. Tranchida, C. Trott, and S. J. Plimpton, LAMMPS - a flexible simulation tool for particle-based materials modeling at the atomic, meso, and continuum scales, Computer Physics Comm...
2022
-
[57]
C. A. Brackley, D. Michieletto, F. Mouvet, J. John- son, S. Kelly, P. R. Cook, and D. Marenduzzo, Simu- lating topological domains in human chromosomes with a fitting-free model, Nucleus7, 453 (2016)
2016
-
[58]
Zhang, D
Y. Zhang, D. S. Lee, Y. Meir, C. P. Brangwynne, and N. S. Wingreen, Mechanical Frustration of Phase Sepa- ration in the Cell Nucleus by Chromatin, Physical Review Letters126, 258102 (2021)
2021
-
[59]
M. M. C. Tortora, L. D. Brennan, G. Karpen, and D. Jost, HP1-driven phase separation recapitulates the thermodynamics and kinetics of heterochromatin con- densate formation, Proceedings of the National Academy of Sciences120, e2211855120 (2023)
2023
-
[60]
J. D. McGhee and G. Felsenfeld, Nucleosome Structure, Annual Review of Biochemistry49, 1115 (1980)
1980
-
[61]
D. E. Olins and A. L. Olins, Chromatin history: our view from the bridge, Nature Reviews Molecular Cell Biology 4, 809 (2003)
2003
-
[62]
J. Widom, A relationship between the helical twist of DNA and the ordered positioning of nucleosomes in all eukaryotic cells., Proceedings of the National Academy of Sciences89, 1095 (1992)
1992
-
[63]
Schalch, S
T. Schalch, S. Duda, D. F. Sargent, and T. J. Richmond, X-ray structure of a tetranucleosome and its implications for the chromatin fibre, Nature436, 138 (2005)
2005
-
[64]
Lieberman-Aiden, N
E. Lieberman-Aiden, N. L. Van Berkum, L. Williams, M. Imakaev, T. Ragoczy, A. Telling, I. Amit, B. R. La- joie, P. J. Sabo, M. O. Dorschner, R. Sandstrom, B. Bern- stein, M. A. Bender, M. Groudine, A. Gnirke, J. Stama- toyannopoulos, L. A. Mirny, E. S. Lander, and J. Dekker, Comprehensive Mapping of Long-Range Interactions Re- veals Folding Principles of ...
2009
-
[65]
Fudenberg, M
G. Fudenberg, M. Imakaev, C. Lu, A. Goloborodko, N. Abdennur, and L. Mirny, Formation of Chromoso- mal Domains by Loop Extrusion, Cell Reports15, 2038 (2016)
2038
-
[66]
J. W. Cahn, Critical point wetting, The Journal of Chem- ical Physics66, 3667 (1977)
1977
-
[67]
P. G. De Gennes, Wetting: statics and dynamics, Re- views of Modern Physics57, 827 (1985)
1985
-
[68]
Binder, D
K. Binder, D. Landau, and M. M¨ uller, Monte Carlo Studies of Wetting, Interface Localization and Capillary Condensation, Journal of Statistical Physics110, 1411 (2003)
2003
-
[69]
J. A. Morin, S. Wittmann, S. Choubey, A. Klosin, S. Golfier, A. A. Hyman, F. J¨ ulicher, and S. W. Grill, Surface condensation of a pioneer transcription factor on DNA (2020)
2020
-
[70]
Quail, S
T. Quail, S. Golfier, M. Elsner, K. Ishihara, V. Muruge- san, R. Renger, F. J¨ ulicher, and J. Brugu´ es, Force gen- eration by protein-DNA co-condensation (2020)
2020
-
[71]
E. E. Alemasova and O. I. Lavrik, Poly(ADP-ribose) in Condensates: The PARtnership of Phase Separation and Site-Specific Interactions, International Journal of Molec- ular Sciences23, 14075 (2022)
2022
-
[72]
Mazzocca, A
M. Mazzocca, A. Loffreda, E. Colombo, T. Fillot, D. Gnani, P. Falletta, E. Monteleone, S. Capozi, E. Bertrand, G. Legube, Z. Lavagnino, C. Tacchetti, and D. Mazza, Chromatin organization drives the search mechanism of nuclear factors, Nature Communications 14, 6433 (2023)
2023
-
[73]
J. A. Morin, S. Wittmann, S. Choubey, A. Klosin, S. Golfier, A. A. Hyman, F. J¨ ulicher, and S. W. Grill, Sequence-dependent surface condensation of a pioneer transcription factor on DNA, Nature Physics18, 271 (2022)
2022
-
[74]
Garc´ ıa Fern´ andez, J
F. Garc´ ıa Fern´ andez, J. Park, C. Chapuis, E. Pinto Ju- rado, V. Imburchia, R. Smith, E. Jos´ e Longarini, A. Tad- dei, C. Hubert, N. Sokolovska, I. Mati´ c, S. Huet, and J. Min´ e-Hattab, Single nucleosome imaging reveals prin- ciples of transient multiscale chromatin reorganization triggered by histone ADP-ribosylation at DNA lesions, Nature Communic...
2025
-
[75]
B. R. Sabari, A. Dall’Agnese, A. Boija, I. A. Klein, E. L. Coffey, K. Shrinivas, B. J. Abraham, N. M. Hannett, A. V. Zamudio, J. C. Manteiga, C. H. Li, Y. E. Guo, D. S. Day, J. Schuijers, E. Vasile, S. Malik, D. Hnisz, T. I. Lee, I. I. Cisse, R. G. Roeder, P. A. Sharp, A. K. Chakraborty, and R. A. Young, Coactivator condensa- tion at super-enhancers links...
2018
-
[76]
Cho, J.-H
W.-K. Cho, J.-H. Spille, M. Hecht, C. Lee, C. Li, V. Grube, and I. I. Cisse, Mediator and RNA polymerase II clusters associate in transcription-dependent conden- sates, Science361, 412 (2018)
2018
-
[77]
Lindahl and D
T. Lindahl and D. Barnes, Repair of Endogenous DNA Damage, Cold Spring Harbor Symposia on Quantitative Biology65, 127 (2000)
2000
-
[78]
Min´ e-Hattab, S
J. Min´ e-Hattab, S. Liu, and A. Taddei, Repair Foci as Liquid Phase Separation: Evidence and Limitations, Genes13, 1846 (2022)
2022
-
[79]
Fries, J
J. Fries, J. Diaz, M. Jardat, I. Pagonabarraga, P. Il- lien, and V. Dahirel, Active droplets controlled by enzy- matic reactions, Journal of The Royal Society Interface 22, 20240803 (2025)
2025
-
[80]
Berthin, J
R. Berthin, J. D. Fries, M. Jardat, V. Dahirel, and P. Il- lien, Microscopic and stochastic simulations of chemically active droplets, Physical Review E111, L023403 (2025)
2025
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.