Recognition: 2 theorem links
· Lean TheoremMultiscale order, flocking and phenotypic hysteresis in the cellular Potts model of epithelia
Pith reviewed 2026-05-15 03:08 UTC · model grok-4.3
The pith
In the cellular Potts model of epithelia, increasing actin polymerization rate drives a transition to long-range flocking with multiscale orientational order and phenotypic hysteresis.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
By accounting for both cell morphology and cytoskeletal activity, the cellular Potts model simulations uncover a remarkably rich phase diagram featuring multiple types of orientational order, either as distinct phases or coexisting across length scales. A gradual increase in the actin polymerization rate drives a phase transition into a long-range flocking state. Simultaneously, quasi-long-range nematic order emerges at length scales much larger than the cell size due to the combined effects of directed motion and lateral cell-cell interactions. At length scales comparable to cell size, however, cells adopt an approximately hexagonal morphology, resulting in hexanematic order. With further增加
What carries the argument
The cellular Potts model that includes explicit cell morphology and cytoskeletal activity controlled by the actin polymerization rate as the tuning parameter.
If this is right
- Increasing the actin polymerization rate along a defined pathway produces a transition into a long-range flocking state.
- Quasi-long-range nematic order develops at scales much larger than individual cells from directed motion combined with lateral interactions.
- Hexanematic order forms at cell-size scales because cells adopt approximately hexagonal shapes.
- At higher polymerization rates nematic order becomes fully long-range while hexatic order remains quasi-long-range and confined to short scales.
- Cycling the monolayer across the melting transition at sufficiently low noise produces phenotypic hysteresis.
Where Pith is reading between the lines
- Real epithelial tissues may achieve different collective behaviors by modulating actin activity to select among the model's ordered states.
- The competition between cell shape and self-propulsion that generates multiscale order could appear in other active-matter systems with deformable particles.
- Experiments could test the model by varying actin regulators in cultured monolayers while tracking order at cell and tissue scales.
- The observed hysteresis offers a possible mechanism for tissues to retain memory of past activity changes after conditions return to normal.
Load-bearing premise
The cellular Potts model's specific rules for cell adhesion, volume constraint, and actin-driven motility capture the essential physics of real epithelial monolayers.
What would settle it
Direct measurement of order parameters in MDCK monolayers that either confirms or rules out the predicted transition to long-range flocking together with the specific multiscale nematic and hexatic orders when actin polymerization rate is raised.
Figures

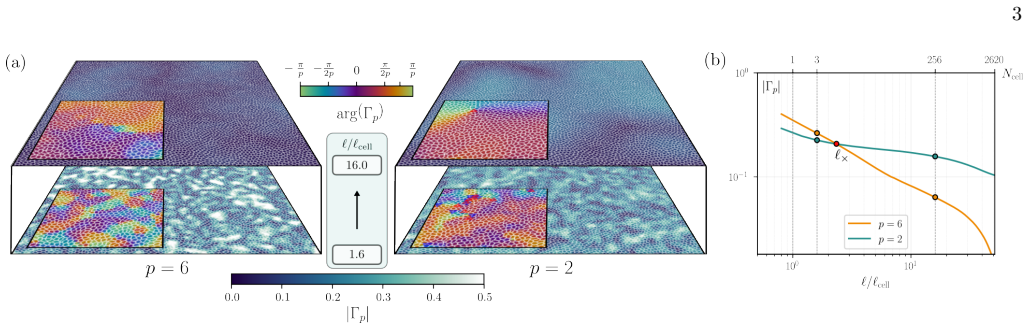


read the original abstract
In epithelia, how do collective cell migration and tissue spatial organization feedback on each other? We address this question through large-scale numerical simulations of the cellular Potts model. By accounting for both cell morphology and cytoskeletal activity, we uncover a remarkably rich phase diagram featuring multiple types of orientational order, either as distinct phases or coexisting across length scales. We identify a specific pathway in parameter space along which a gradual increase in the actin polymerization rate drives a phase transition into a long-range flocking state. Simultaneously, quasi-long-range nematic order emerges at length scales much larger than the cell size due to the combined effects of directed motion and lateral cell-cell interactions. At length scales comparible to cell size, however, cells adopt an approximatively hexagonal morphology, resulting in hexanematic order, similar to that observed in reconstituted Madin-Darby Canine Kidney (MDCK) cell monolayers. With further increases in actin polymerization, nematic order becomes fully long-range, while hexatic order remains quasi-long-range and confined to short length scales, but independent of cytoskeletal activity. When noise is sufficiently low to allow crystallization at finite actin polymerization rate, cycling the cell-monolayer across the melting transition yields an example of phenotypical hysteresis, reminiscent of that observed across the epithelial-mesenchymal transition.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript uses large-scale cellular Potts model simulations that incorporate cell morphology and cytoskeletal activity to investigate feedback between collective migration and spatial organization in epithelia. It reports a rich phase diagram containing multiple orientational orders (distinct or coexisting across scales), identifies a parameter pathway in which increasing actin polymerization rate drives a transition into a long-range flocking state, describes emergence of quasi-long-range nematic order at large scales and hexanematic order at cell scales, and demonstrates phenotypic hysteresis upon cycling across a melting transition at low noise.
Significance. If the central claims on long-range flocking and multiscale order are confirmed, the work would be significant for linking cytoskeletal parameters to tissue-level collective behaviors and for explaining experimental observations such as hexanematic order in MDCK monolayers. The use of an established CPM framework to explore scale-dependent orders and hysteresis is a strength, as is the identification of a specific, gradual parameter pathway for the flocking transition. The results could inform models of epithelial-mesenchymal transitions if the orders are shown to be robust beyond finite-size effects.
major comments (2)
- [Abstract] Abstract: The claim of a transition into a 'long-range flocking state' and 'quasi-long-range nematic order' at scales much larger than the cell size requires finite-size scaling analysis of the relevant order parameters (polarization magnitude and nematic tensor) with linear system size L. In 2D systems with continuous rotational symmetry the Mermin-Wagner theorem precludes true long-range order, so it is essential to demonstrate that the polarization does not decay with L and that the structure-factor peak remains finite in the thermodynamic limit rather than arising from correlation lengths comparable to the simulated box size.
- [Abstract] Abstract: Simulation details necessary to assess the reported orders are absent, including the lattice sizes (or number of cells) employed, the precise definitions and normalizations of the order parameters used to quantify flocking, nematic, and hexatic order, and any scaling collapses or finite-size extrapolations performed. Without these, it is impossible to determine whether the claimed long-range flocking is an artifact of the finite systems or a genuine thermodynamic feature.
minor comments (2)
- [Abstract] Typographical error: 'comparible' should read 'comparable'.
- [Abstract] Typographical error: 'approximatively' should read 'approximately'.
Simulated Author's Rebuttal
We thank the referee for their careful reading and constructive feedback on our manuscript. We address the major comments point by point below. We agree that additional details and analyses are needed to fully substantiate the claims and will revise the manuscript accordingly.
read point-by-point responses
-
Referee: [Abstract] Abstract: The claim of a transition into a 'long-range flocking state' and 'quasi-long-range nematic order' at scales much larger than the cell size requires finite-size scaling analysis of the relevant order parameters (polarization magnitude and nematic tensor) with linear system size L. In 2D systems with continuous rotational symmetry the Mermin-Wagner theorem precludes true long-range order, so it is essential to demonstrate that the polarization does not decay with L and that the structure-factor peak remains finite in the thermodynamic limit rather than arising from correlation lengths comparable to the simulated box size.
Authors: We agree that finite-size scaling is essential to rigorously support claims of long-range or quasi-long-range order in two-dimensional systems, given the Mermin-Wagner theorem. In the revised manuscript we will add a dedicated finite-size scaling analysis of the polarization magnitude (flocking order parameter) and the nematic tensor as functions of linear system size L. We will show that, along the identified parameter pathway, the flocking polarization remains finite and does not decay with increasing L (up to the largest simulated sizes), while the nematic order parameter exhibits the algebraic decay characteristic of quasi-long-range order. These results will be presented with appropriate scaling plots to confirm that the observed orders are not artifacts of finite correlation lengths comparable to the box size. revision: yes
-
Referee: [Abstract] Abstract: Simulation details necessary to assess the reported orders are absent, including the lattice sizes (or number of cells) employed, the precise definitions and normalizations of the order parameters used to quantify flocking, nematic, and hexatic order, and any scaling collapses or finite-size extrapolations performed. Without these, it is impossible to determine whether the claimed long-range flocking is an artifact of the finite systems or a genuine thermodynamic feature.
Authors: We apologize for the omission of these methodological details from the abstract. The full manuscript already specifies the lattice sizes (up to 512×512 sites, corresponding to several thousand cells) and the definitions of the order parameters: the flocking polarization is the magnitude of the spatially averaged cell velocity vector; the nematic order parameter is obtained from the traceless symmetric tensor constructed from cell elongation axes; and the hexatic order parameter follows the standard six-fold bond-orientational definition. In the revision we will add an explicit Methods subsection (or appendix) that provides the precise mathematical definitions, normalizations, and the finite-size scaling collapses already performed. This will enable readers to independently verify the robustness of the reported phases. revision: yes
Circularity Check
No significant circularity in simulation-based exploration
full rationale
The paper reports results from large-scale numerical simulations of the established cellular Potts model, varying parameters such as actin polymerization rate to map a phase diagram with orientational orders and flocking. No analytical derivation chain exists in which a claimed prediction or first-principles result reduces by construction to its inputs, fitted parameters, or self-citations. Behaviors such as hexanematic order and hysteresis emerge dynamically from the model rules rather than being tautologically defined. The central claims rest on computational outcomes, not self-referential definitions or load-bearing self-citations that collapse the argument.
Axiom & Free-Parameter Ledger
free parameters (2)
- actin polymerization rate
- noise level
axioms (2)
- domain assumption The cellular Potts model with morphology and cytoskeletal activity terms accurately models epithelial dynamics
- domain assumption Cell-cell interactions and directed motion lead to observed orders
Lean theorems connected to this paper
-
IndisputableMonolith/Foundation/AlexanderDuality.leanalexander_duality_circle_linking unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
large-scale numerical simulations of the cellular Potts model... Hamiltonian H = J Σ(1−δ_σiσj) + λ_cell Σ(Ac−A0)² + λ_cyto Σ GM(ϕi/Φ)
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
phase diagram... hexanematic order... long-range flocking state... η_p exponents
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
J.-M. Armengol-Collado, L. N. Carenza, J. Eckert, D. Krommydas, and L. Giomi, Nat. Phys.19, 1773 (2023)
work page 2023
- [2]
-
[3]
J.-M. Armengol-Collado, L. N. Nicola, and L. Giomi, eLife13, e86400 (2024)
work page 2024
-
[4]
L. N. Carenza, J.-M. Armengol-Collado, D. Krommydas, and L. Giomi, Phys. Rev. Lett.134, 128304 (2025)
work page 2025
- [5]
- [6]
-
[7]
Y.-W. Pasupalak, A.and Li, R. Ni, and M. Pica Cia- marra, Soft Matter16, 3914 (2020)
work page 2020
- [8]
- [9]
-
[10]
B. I. Halperin and D. R. Nelson, Phys. Rev. Lett.41, 121 (1978)
work page 1978
-
[11]
D. R. Nelson and B. I. Halperin, Phys. Rev. B19, 2457 (1979)
work page 1979
-
[12]
A. P. Young, Phys. Rev. B19, 1855 (1979)
work page 1979
-
[13]
C. Carmona-Fontaine, H. K. Matthews, S. Kuriyama, M. Moreno, G. A. Dunn, M. Parsons, J. D. Stern, and R. Mayor, Nature456, 957 (2008)
work page 2008
- [14]
-
[15]
C. Malinverno, S. Corallino, F. Giavazzi, M. Bergert, Q. Li, M. Leoni, A. Disanza, E. Frittoli, A. Oldani, E. Martini, T. Lendenmann, G. Deflorian, G. V. Beznoussenko, D. Poulikakos, O. K. Haur, M. Uroz, X. Trepat, D. Parazzoli, P. Maiuri, W. Yu, A. Ferrari, R. Cerbino, and G. Scita, Nat. Mater.16, 587 (2017)
work page 2017
-
[16]
F. Giavazzi, C. Malinverno, S. Corallino, F. Ginelli, G. Scita, and R. Cerbino, J. Phys. D: Appl. Phys.50, 474003 (2017)
work page 2017
-
[17]
A. Glentis, C. Blanch-Mercader, L. Balasubramaniam, T. B. Saw, J. d’Alessandro, D. Delacour, J. Prost, W. Xi, and B. Ladoux, Sci. Adv.8, eabn5406 (2022)
work page 2022
- [18]
- [19]
-
[20]
A. Carrillo, M. S. Figueroa, P. J. Davis, A. E. Aguilar, T. Hines, M. L. Van Avermaete, D. B. Zhang, C. A. Alvarado, P. E. Kuo, A. Kim, G. L. Quintero, P. F. Auer, M. J. Benskey, M. G. Castro, and P. R. Lowenstein, Nat. Commun.13, 3606 (2022)
work page 2022
-
[21]
A. E. Argento, M. L. Varela, G. Singh, D. P. Vis- nuk, B. Jacobovitz, M. E. Rutherford, M. B. Edwards, Q. Chaboche, D. A. Orringer, J. A. Heth, M. G. Castro, D. A. Beller, C. Blanch-Mercader, and P. R. Lowenstein, bioRxiv 10.1101/2025.04.14.648711 (2025)
-
[22]
C. J. Weijer, J. Cell Sci.122, 3215 (2009)
work page 2009
- [23]
-
[24]
F. Giavazzi, M. Paoluzzi, M. Macchi, D. Bi, G. Scita, M. L. Manning, R. Cerbino, and M. C. Marchetti, Soft Matter14, 3471 (2018)
work page 2018
-
[25]
P. Baconnier, O. Dauchot, V. D´ emery, G. D¨ uring, S. Henkes, C. Huepe, and A. Shee, Rev. Mod. Phys.97, 015007 (2025)
work page 2025
- [26]
-
[27]
P. Yu, F. Brauns, and M. C. Marchetti, preprint arXiv:2511.04189 (2025)
work page internal anchor Pith review Pith/arXiv arXiv 2025
- [28]
-
[29]
J. A. Glazier and F. Graner, Physical Review E47, 2128 (1993)
work page 1993
- [30]
-
[31]
I. Niculescu, J. Textor, and R. J. De Boer, PLoS comput. biol.11, e1004280 (2015)
work page 2015
-
[32]
I. M. Wortel, I. Niculescu, P. M. Kolijn, N. S. Gov, R. J. de Boer, and J. Textor, Biophys. J.120, 2609 (2021)
work page 2021
-
[33]
L. van Steijn, I. M. Wortel, C. Sire, L. Dupr´ e, G. Ther- aulaz, and R. M. Merks, PLoS Comput. Biol.18, e1009156 (2022)
work page 2022
-
[34]
T. Toffoli and N. Margolus,Cellular automata machines: a new environment for modeling(MIT Press, Cambridge, MA, USA, 1987)
work page 1987
-
[35]
A MCS here is equal to an amount of update-attempts of the Metropolis-Hastings algorithm that is equal to the amount of pixels in the two-dimensionalL×Larray:L 2
- [36]
-
[37]
Supplementary information, inclusive of Refs. [51–57]
- [38]
- [39]
- [40]
-
[41]
T. K. Ho, inProc. Int. Conf. Doc. Anal. Recognit., Vol. 1 (IEEE, 1995) pp. 278–282
work page 1995
-
[42]
W. E. Lorensen and H. E. Cline, inSeminal Graphics: Pioneering Efforts That Shaped the Field(ACM, 1998) pp. 347–353
work page 1998
-
[43]
Here the hexatetratic phase and more exotic phases were excluded from the visualization (see Ref. [37] for details)
- [44]
-
[45]
J. V. Selinger and D. R. Nelson, Phys. Rev. A39, 3135 (1989)
work page 1989
-
[46]
J. V. Selinger, J. Phys. II (France)1, 1363 (1991)
work page 1991
-
[47]
V. Drouin-Touchette, P. P. Orth, P. Coleman, P. Chan- dra, and T. C. Lubensky, Phys. Rev. X12, 011043 (2022)
work page 2022
- [48]
-
[49]
K. K. Youssef, N. Narwade, A. Arcas, A. Marquez- Galera, R. Jim´ enez-Casta˜ no, C. Lopez-Blau, H. Fazilaty, D. Garc´ ıa-Gutierrez, A. Cano, J. Galcer´ an, G. Moreno- Bueno, J. P. Lopez-Atalaya, and M. A. Nieto, Nat. Can- cer5, 1660 (2024)
work page 2024
-
[50]
A. S. Jahun, F. Sorgeloos, Y. Chaudhry, S. E. Arthur, M. Hosmillo, I. Georgana, R. Izuagbe, and I. G. Good- fellow, Cell Rep.42, 112179 (2023)
work page 2023
- [51]
-
[52]
F. Pedregosa, G. Varoquaux, A. Gramfort, V. Michel, B. Thirion, O. Grisel, M. Blondel, P. Prettenhofer, R. Weiss, V. Dubourg, J. Vanderplas, A. Passos, D. Cour- napeau, M. Brucher, M. Perrot, and ´E. Duchesnay, J. Mach. Learn. Res.12, 2825 (2011)
work page 2011
-
[53]
S. van der Walt, J. L. Sch¨ onberger, J. N´ u˜ nez-Iglesias, F. Boulogne, J. D. Warner, N. Yager, E. Gouillart, and T. Yu, PeerJ2, e453 (2014)
work page 2014
-
[54]
D. Bi, X. Yang, M. C. Marchetti, and M. L. Manning, Phys. Rev. X6, 021011 (2016)
work page 2016
- [55]
- [56]
-
[57]
On the Concept of Static Structure Factor
K. Zhang, preprint arXiv:1606.03610 (2016). Multiscale order, flocking and phenotypic hysteresis in the cellular Potts model of epithelia: Supplementary information Calvin C. Bakker, 1 Marc Durand, 2 Fran¸ cois Graner,2 and Luca Giomi 1 1Instituut-Lorentz, Leiden Institute of Physics, Universiteit Leiden, P.O. Box 9506, 2300 RA Leiden, The Netherlands 2Un...
work page internal anchor Pith review Pith/arXiv arXiv 2016
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.