PACE: Geometry-Aware Bridge Transport for Single-Cell Trajectory Inference
Pith reviewed 2026-05-20 01:20 UTC · model grok-4.3
The pith
PACE recovers continuous cell trajectories from snapshots by building a time-varying anisotropic metric that favors local developmental directions.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
PACE shows that a state- and time-dependent anisotropic Riemannian metric can be used to define path-action costs that enforce geometry-consistent couplings between snapshots; alternating optimization between these costs and neural bridge fitting then produces a distilled global velocity field that reconstructs held-out trajectories more accurately than Euclidean optimal transport or flow-based baselines.
What carries the argument
A state- and time-dependent anisotropic Riemannian metric that assigns low transport cost along locally supported tangent directions while penalizing normal velocity components.
If this is right
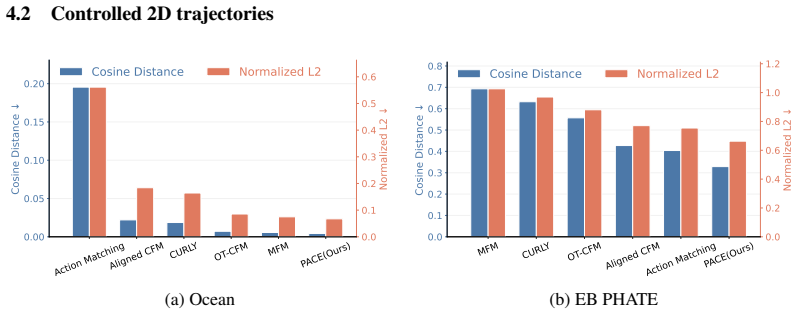
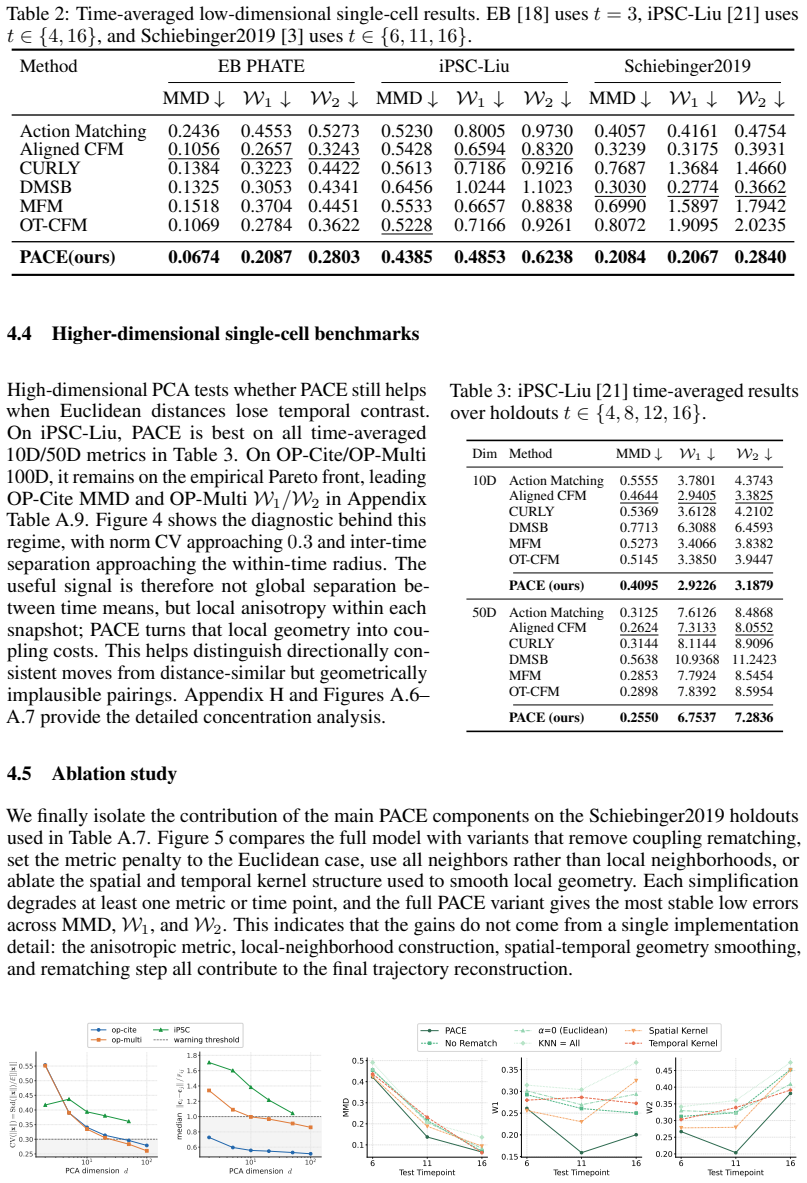
- Reduces MMD, Wasserstein-1, and Wasserstein-2 distances by 23.7 percent on average across nine reconstruction experiments on seven datasets.
- Improves alignment with measured RNA velocity by 15.4 percent on an embryoid body differentiation benchmark.
- Recovers continuous dynamics without requiring explicit cell pairing, lineage tracing, or RNA-velocity supervision during training.
Where Pith is reading between the lines
- The same metric construction could be tested on other asynchronous dynamical systems outside single-cell biology where local geometry is known to matter.
- Datasets that supply ground-truth continuous trajectories would allow direct measurement of how much the anisotropy assumption improves path accuracy versus endpoint matching alone.
- Combining the distilled velocity field with multi-omics measurements could yield joint trajectory models across transcriptomic and proteomic layers.
Load-bearing premise
That a suitable anisotropic metric reflecting local tangent directions of development can be constructed from the observed snapshots alone.
What would settle it
A controlled simulation in which true trajectories follow known curved paths but the learned metric forces straighter couplings would produce higher reconstruction errors than Euclidean baselines on the same data.
Figures


read the original abstract
Single-cell trajectory inference from destructive time-course snapshots is fundamentally ill-posed: neither cross-time cell correspondences nor continuous trajectories are observed, so the snapshot distributions alone do not uniquely determine the underlying dynamics. Existing optimal transport and flow-based methods typically couple cells by Euclidean proximity at observed clock times, which can misalign trajectories when development is asynchronous and cells sampled at the same experimental time occupy different latent pseudotime stages. We propose PACE, a trajectory inference framework that recovers geometry-consistent continuous transport dynamics from destructive time-course snapshots through three coupled components. First, PACE constructs a state- and time-dependent anisotropic Riemannian metric that assigns low transport cost along locally supported tangent directions while penalizing normal velocity components. Second, it alternates between refining cross-time couplings under the induced path-action cost and fitting endpoint-preserving neural bridges between adjacent snapshots. Third, it distills the learned bridge dynamics into a global continuous-time velocity field over cellular states. Across seven controlled and biological datasets covering nine held-out reconstruction experiments, PACE achieves the strongest overall reconstruction performance, reducing MMD, Wasserstein-1 distance, and Wasserstein-2 distance by 23.7% on average relative to the strongest competing baseline. PACE also improves RNA-velocity alignment by 15.4% on an embryoid body differentiation benchmark, without requiring explicit cell pairing, lineage tracing, or RNA-velocity supervision during training. Code is available at https://github.com/AI4Science-WestlakeU/PACE.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper proposes PACE, a trajectory inference method for single-cell data from destructive time-course snapshots. It constructs a state- and time-dependent anisotropic Riemannian metric that favors transport along locally supported tangent directions, alternates between optimizing cross-time couplings under the induced path cost and fitting endpoint-preserving neural bridges, and distills the dynamics into a global continuous-time velocity field. On seven controlled and biological datasets with nine held-out reconstruction experiments, it reports a 23.7% average reduction in MMD, Wasserstein-1, and Wasserstein-2 distances relative to the strongest baseline, plus a 15.4% improvement in RNA-velocity alignment on an embryoid-body benchmark, without requiring cell pairing or velocity supervision.
Significance. If the central geometry-consistency claim holds, PACE would address a key limitation of Euclidean OT and flow-based methods in asynchronous developmental settings by penalizing normal velocity components. The open-source code at the provided GitHub link is a clear strength for reproducibility and further testing. The empirical gains on held-out reconstruction tasks suggest practical utility, but only if the performance can be attributed to the Riemannian metric rather than the neural-bridge or distillation components alone.
major comments (2)
- [Abstract and §3] Abstract and §3 (metric construction): the headline 23.7% average improvement is load-bearing for the geometry-awareness claim, yet the manuscript provides no explicit formula, algorithm, or pseudocode for estimating local tangent directions from snapshots (e.g., via local PCA or similar). Without this, it is impossible to verify that the anisotropic metric reliably penalizes normal components rather than reverting to near-Euclidean behavior under noise.
- [§4 and Table 2] §4 (experiments) and Table 2: the nine held-out reconstruction experiments report average percentage reductions but omit per-experiment error bars, standard deviations across random seeds, and an ablation that isolates the Riemannian metric from the neural-bridge fitting and distillation steps. This prevents assessment of whether the reported superiority is robust or driven by the geometry component.
minor comments (2)
- [§2] Notation for the path-action cost and the time-dependent metric tensor is introduced without a clear summary table relating symbols to their definitions, which would aid readability.
- [§4.3] The RNA-velocity alignment experiment on the embryoid-body benchmark is described only in the abstract; a dedicated subsection with the exact alignment metric and baseline details would strengthen the biological validation.
Simulated Author's Rebuttal
We thank the referee for the thoughtful and constructive comments on our manuscript. We address each of the major comments below and have revised the manuscript to incorporate the suggested improvements for greater clarity and rigor.
read point-by-point responses
-
Referee: [Abstract and §3] Abstract and §3 (metric construction): the headline 23.7% average improvement is load-bearing for the geometry-awareness claim, yet the manuscript provides no explicit formula, algorithm, or pseudocode for estimating local tangent directions from snapshots (e.g., via local PCA or similar). Without this, it is impossible to verify that the anisotropic metric reliably penalizes normal components rather than reverting to near-Euclidean behavior under noise.
Authors: We appreciate this observation. Section 3 describes the use of local PCA on cell neighborhoods within each snapshot to approximate tangent directions for the state- and time-dependent anisotropic metric. To improve verifiability and address the concern directly, we have added explicit pseudocode and the precise mathematical formula for the metric tensor construction in a new algorithm box in the revised Section 3. This addition clarifies how normal components are penalized and allows readers to assess behavior under noise. revision: yes
-
Referee: [§4 and Table 2] §4 (experiments) and Table 2: the nine held-out reconstruction experiments report average percentage reductions but omit per-experiment error bars, standard deviations across random seeds, and an ablation that isolates the Riemannian metric from the neural-bridge fitting and distillation steps. This prevents assessment of whether the reported superiority is robust or driven by the geometry component.
Authors: We agree that these details are important for assessing robustness. In the revised manuscript, we have updated Table 2 to report per-experiment means with error bars and standard deviations across five random seeds. We have also added an ablation study in Section 4.3 (with corresponding results in the supplement) that isolates the Riemannian metric by comparing full PACE against variants that disable the anisotropic component while retaining the neural-bridge fitting and distillation steps. The ablation confirms the geometry component drives a substantial portion of the gains. revision: yes
Circularity Check
No significant circularity detected in derivation chain
full rationale
The provided abstract and description outline a three-component framework: construction of a state- and time-dependent anisotropic Riemannian metric, alternation between cross-time couplings and neural bridge fitting, and distillation to a global velocity field. Performance is evaluated empirically via MMD, Wasserstein-1, and Wasserstein-2 distances against external baselines on held-out experiments, with no equations or self-referential definitions that reduce claimed gains to inputs by construction. No load-bearing self-citations, fitted parameters renamed as predictions, or ansatzes smuggled via prior work appear in the text. The chain remains self-contained with independent empirical support.
Axiom & Free-Parameter Ledger
free parameters (2)
- neural bridge network parameters
- metric anisotropy parameters
axioms (1)
- domain assumption A state- and time-dependent anisotropic Riemannian metric exists that correctly captures locally supported tangent directions for cellular transport.
Lean theorems connected to this paper
-
IndisputableMonolith/Foundation/AlexanderDuality.leanalexander_duality_circle_linking unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
PACE constructs a state- and time-dependent anisotropic Riemannian metric that assigns low transport cost along locally supported tangent directions while penalizing normal velocity components... Local PCA... Gaussian-kernel weighted covariance... normal projector P_N^{(r)}
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
The metric turns the qualitative principle of geometry-aligned cellular motion into an explicit action functional... v⊤ G v = ||v_T||² + (1+α)||v_N||²
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.