Rate-induced tipping in a coral reef ecosystem: A slow increase in fishing effort can induce reef collapse
Pith reviewed 2026-05-25 02:52 UTC · model grok-4.3
The pith
A slow rise in fishing effort can trigger sudden coral reef collapse through rate-induced tipping.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
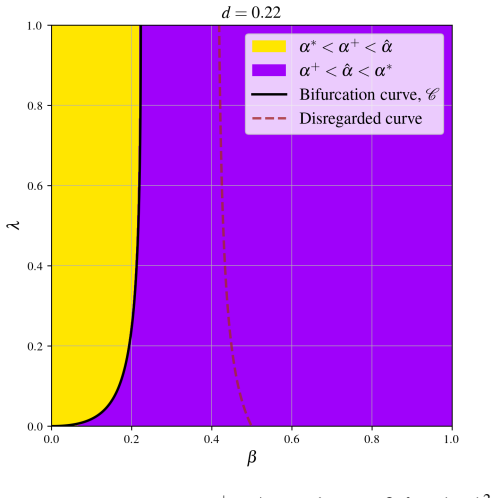
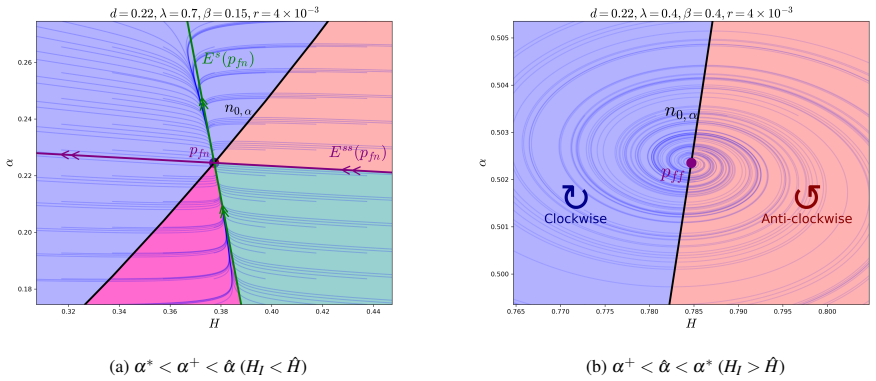
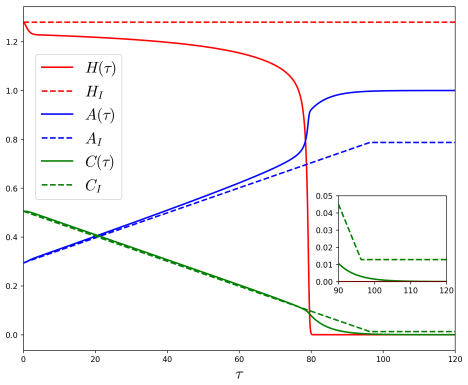
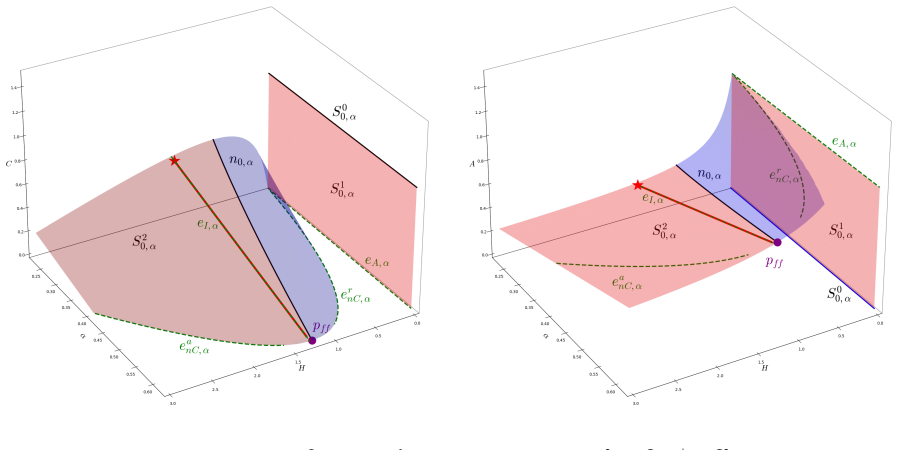
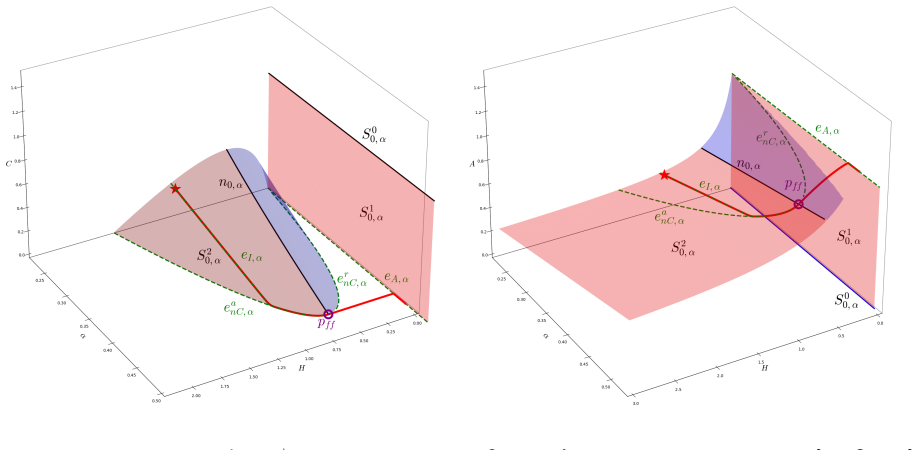
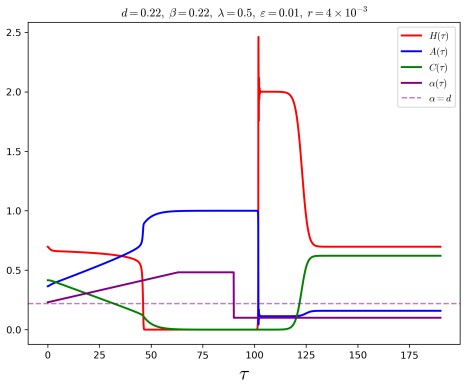
In the behavioural-demographic model for herbivorous fish, algae and coral, rate-induced tipping occurs when fishing effort increases at a rate too fast for the ecosystem to adapt; trajectories undergo canard-induced tipping by passage through a folded node singularity or jump-type tipping through a folded focus, in both cases producing catastrophic collapse of fish and coral populations together with an algae bloom, while tracking of the coexistence state remains possible only for certain folded-focus regimes.
What carries the argument
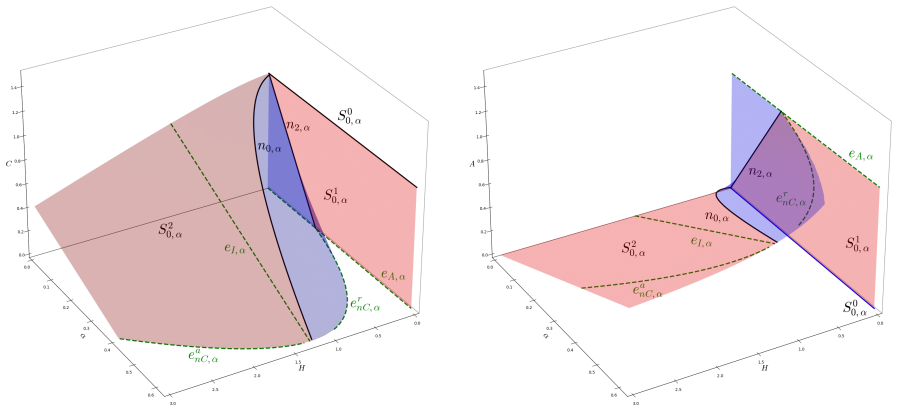
The singularly perturbed system with two fast variables and one slow variable, analysed via geometric singular perturbation theory, in which folded node and folded focus singularities determine whether trajectories tip or track.
If this is right
- Slow but sustained increases in fishing effort produce sudden population collapses instead of smooth adaptation.
- Canard-induced tipping through a folded node yields one class of collapse while passage near a folded focus yields a jump-type collapse.
- Algae populations exhibit a bloom precisely when fish and coral collapse under either tipping mechanism.
- Tracking of the sustainable three-species coexistence state is possible only in parameter regimes containing a folded focus.
Where Pith is reading between the lines
- Management strategies may need to track the rate of change in fishing pressure, not only its absolute level, to avoid crossing tipping thresholds.
- The same rate-induced mechanism could appear in other multi-species models whenever a slow anthropogenic driver acts on a fast ecological subsystem.
- Numerical continuation or field measurements of population trajectories near candidate folded singularities could locate the critical rates of effort increase.
Load-bearing premise
The demographic model can be reframed as a singularly perturbed system that admits bistability in ecologically relevant parameter regimes.
What would settle it
A controlled increase in fishing effort at a constant slow rate that produces only gradual declines in fish and coral without an abrupt collapse or algae bloom would falsify the predicted tipping.
Figures














read the original abstract
Critical transitions describe sudden changes in the state of an ecosystem. In classical bifurcation theory, such transitions occur when the value of a parameter exceeds a threshold (``bifurcation") value. More recently, critical transitions which are triggered by the rate of change of a parameter were described by Wieczorek et al. [Wieczorek, S., Ashwin, P., Luke, C.M., Cox, P.M., Proceedings of the Royal Society A 467(2129), 1243-1269, 2011]. In mathematical ecology, these rate-induced transitions correspond to environmental conditions that deteriorate too rapidly for the ecosystem to adapt, resulting in population collapse (``R-tipping"). In this article, we consider the potential for rate-induced tipping due to increased anthropogenic stress in a recently proposed behavioural-demographic model for herbivorous fish, algae, and coral in a coral reef ecosystem [Gil, M.A., Baskett, M.L., Munch, S.B., Hein, A.M., PNAS 117(41), 25580-25589, 2020]. We first show that the underlying demographic model can be reframed naturally as a singularly perturbed system with two fast variables and one slow variable in which bistability can occur in ecologically relevant parameter regimes. We explore the potential for canard-type dynamics in the model, complementing numerical results with an analytical description through the lens of geometric singular perturbation theory, and we describe R-tipping as a result of an increase in the fishing effort. We show that trajectories will undergo canard-induced tipping by passage through a folded node singularity, whereas a folded focus may give rise to tipping of jump type; in both scenarios, a catastrophic collapse occurs in the populations of herbivorous fish and coral, with the population of algae experiencing a ``bloom". Alternatively, we may observe ``tracking" of a sustainable coexistence state between the three populations in the presence of a folded focus.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript reframes the Gil et al. (2020) coral-reef demographic model as a 2-fast/1-slow singularly perturbed system and applies geometric singular perturbation theory to show that a slow increase in fishing effort can produce rate-induced tipping. Trajectories either undergo canard-induced tipping by passage through a folded-node singularity or jump-type tipping through a folded focus; both routes produce collapse of herbivorous fish and coral with an algae bloom, while a folded focus can alternatively permit tracking of a sustainable coexistence state.
Significance. If the central claims hold, the work supplies an analytically grounded example of rate-induced tipping in an applied ecological model, distinguishing canard versus jump mechanisms via the geometry of the critical manifold and folded singularities. The combination of GSPT analysis with numerical verification and the explicit link to an existing, parameterised reef model constitute a clear strength.
minor comments (2)
- [Abstract] Abstract, first paragraph after abstract: the statement that 'bistability can occur in ecologically relevant parameter regimes' is central to the subsequent GSPT analysis; a short explicit statement of the parameter ranges (or a reference to the section containing them) would make the claim immediately verifiable.
- The manuscript cites Wieczorek et al. (2011) for the general R-tipping framework; a brief sentence clarifying which of their results are invoked verbatim versus which are adapted to the present 2-fast/1-slow setting would aid readers.
Simulated Author's Rebuttal
We thank the referee for their positive assessment of our manuscript and for recommending minor revision. The summary accurately captures the main contributions regarding the reframing of the Gil et al. (2020) model as a singularly perturbed system and the analysis of rate-induced tipping via canards and folded singularities. No major comments requiring specific responses were provided in the report.
Circularity Check
No significant circularity identified
full rationale
The derivation reframes the externally cited Gil et al. (2020) demographic model as a 2-fast 1-slow singularly perturbed system and applies standard geometric singular perturbation theory to locate folded node/focus singularities and describe canard-induced or jump-type R-tipping. No step reduces a claimed prediction or uniqueness result to a quantity defined or fitted inside this paper; the bistability and tipping outcomes are direct consequences of the cited model's structure under GSPT, with no self-citation chains or ansatz smuggling. The analysis is therefore self-contained against external benchmarks.
Axiom & Free-Parameter Ledger
free parameters (1)
- rate of fishing effort increase
axioms (2)
- domain assumption The demographic model can be reframed naturally as a singularly perturbed system with two fast variables and one slow variable
- domain assumption Bistability can occur in ecologically relevant parameter regimes
Reference graph
Works this paper leans on
-
[1]
Peter Ashwin, Sebastian Wieczorek, Renato Vitolo, and Peter Timothy Cox,Tipping points in open systems: bifurcation, noise-induced and rate-dependent examples in the climate system, Philosophical Transactions of the Royal Society A370(2012), 1166–1184
work page 2012
-
[2]
Blackwood, Alan Hastings, and Peter J
Julie C. Blackwood, Alan Hastings, and Peter J. Mumby,The effect of fishing on hysteresis in caribbean coral reefs, Theoretical Ecology5(2010), 105–114
work page 2010
-
[3]
Cheryl J. Briggs, Thomas C. Adam, Sally J. Holbrook, and Russell J. Schmitt,Macroalgae size refuge from herbivory promotes alternative stable states on coral reefs, PLOS ONE13(2018), e0202273
work page 2018
-
[4]
Peter De Maesschalck, Ekaterina Kutafina, and Nikola Popovi ´c,Sector-delayed-hopf-type mixed-mode oscillations in a prototypical three-time-scale model, Applied Mathematics and Computation273(2016), 337–352
work page 2016
-
[5]
Mathieu Desroches, John Guckenheimer, Bernd Krauskopf, Christian Kuehn, Hinke M. Osinga, and Martin Wechselberger,Mixed-mode oscillations with multiple time scales, SIAM Rev.54(2012), no. 2, 211–288. MR 2916308
work page 2012
-
[6]
Tak Fung, Robert M. Seymour, and Craig R. Johnson,Alternative stable states and phase shifts in coral reefs under anthropogenic stress, Ecology92(2011), 967–982
work page 2011
-
[7]
Michael A. Gil, Marissa L. Baskett, Stephan B. Munch, and Andrew M. Hein,Fast behavioral feedbacks make ecosystems sensitive to pace and not just magnitude of anthropogenic environmental change, Proceedings of the National Academy of Sciences117(2020), no. 41, 25580–25589
work page 2020
-
[8]
Michael A. Gil and Andrew M. Hein,Social interactions among grazing reef fish drive material flux in a coral reef ecosystem, Proceedings of the National Academy of Sciences114(2017), no. 18, 4703–4708
work page 2017
-
[9]
Terence P. Hughes,Catastrophes, phase shifts, and large-scale degradation of a caribbean coral reef, Science 265(1994), 1547–1551
work page 1994
-
[10]
Arieh Iserles,A first course in the numerical analysis of differential equations, second ed., Cambridge Texts in Applied Mathematics, Cambridge University Press, Cambridge, 2009. MR 2478556
work page 2009
-
[11]
Panagiotis Kaklamanos, Nikola Popovi ´c, and Kristian Uldall Kristiansen,Bifurcations of mixed-mode oscillations in three-timescale systems: An extended prototypical example, Chaos32(2022) (English)
work page 2022
-
[12]
Martin Krupa and Peter Szmolyan,Extending geometric singular perturbation theory to nonhyperbolic points—fold and canard points in two dimensions, SIAM Journal on Mathematical Analysis33(2001), no. 2, 286–314
work page 2001
-
[13]
,Extending slow manifolds near transcritical and pitchfork singularities, Nonlinearity14(2001), no. 6, 1473
work page 2001
-
[14]
Christian Kuehn,Multiple time scale dynamics, Applied Mathematical Sciences, vol. 191, Springer, Cham, 2015. 34
work page 2015
-
[15]
Mumby, Alan Hastings, and Helen J
Peter J. Mumby, Alan Hastings, and Helen J. Edwards,Thresholds and the resilience of caribbean coral reefs, Nature450(2007), 98–101
work page 2007
-
[16]
Lawrence F. Shampine and Mark W. Reichelt,The matlab ode suite, SIAM Journal on Scientific Computing18 (1997), no. 1, 1–22
work page 1997
-
[17]
Katharine N. Suding and Richard J. Hobbs,Threshold models in restoration and conservation: a developing framework, Trends in Ecology & Evolution24(2009), 271–279
work page 2009
-
[18]
Peter Szmolyan and Martin Wechselberger,Canards inR 3, Journal of Differential Equations177(2001), no. 2, 419–453
work page 2001
-
[19]
James E. Truscott and John Brindley,Ocean plankton populations as excitable media, Bulletin of Mathematical Biology56(1994), no. 5, 981–998
work page 1994
-
[20]
Anna Vanselow, Lukas Halekotte, Pinaki Pal, Sebastian Wieczorek, and Ulrike Feudel,Rate-induced tipping can trigger plankton blooms, Theoretical Ecology17(2024), 89–105
work page 2024
-
[21]
Anna Vanselow, Sebastian Wieczorek, and Ulrike Feudel,When very slow is too fast - collapse of a predator-prey system, Journal of Theoretical Biology479(2019), 64–72
work page 2019
-
[22]
Pauli Virtanen, Ralf Gommers, Travis E. Oliphant, Matt Haberland, Tyler Reddy, David Cournapeau, Evgeni Burovski, Pearu Peterson, Warren Weckesser, Jonathan Bright, St ´efan J. van der Walt, Matthew Brett, Joshua Wilson, K. Jarrod Millman, Nikolay Mayorov, Andrew R. J. Nelson, Eric Jones, Robert Kern, Eric Larson, C J Carey, ˙Ilhan Polat, Yu Feng, Eric W....
work page 2020
-
[23]
Martin Wechselberger,Existence and bifurcation of canards inR 3 in the case of a folded node, SIAM J. Appl. Dyn. Syst.4(2005), no. 1, 101–139. MR 2136520
work page 2005
-
[24]
, `A propos de canards (Apropos canards), Trans. Amer. Math. Soc.364(2012), no. 6, 3289–3309. MR 2888246
work page 2012
-
[25]
Sebastian Wieczorek, Peter Ashwin, Catherine M. Luke, and Peter M. Cox,Excitability in ramped systems: the compost-bomb instability, Proceedings of the Royal Society A: Mathematical, Physical and Engineering Sciences 467(2011), 1243–1269. 35
work page 2011
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.