CellBRIDGE: Learning Cellular Trajectories via Interaction-Aware Alignment
Pith reviewed 2026-06-29 08:28 UTC · model grok-4.3
The pith
Adding ligand-receptor interaction costs to optimal transport improves alignments and trajectory estimates from cell snapshots.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
CellBRIDGE augments feature-based optimal transport with a directed, typed interaction cost derived from ligand-receptor activity. By explicitly modeling cell-cell communication, CellBRIDGE improves cross-snapshot couplings and downstream trajectory estimates across synthetic and real scRNA-seq datasets relative to feature-only baselines. Notably, CellBRIDGE enables mechanistically interpretable in silico perturbations: on lung cancer data, silencing specific ligand-receptor pairs induces trajectory shifts that recapitulate expected effects of targeted pathway inhibition.
What carries the argument
The interaction-augmented optimal transport cost that adds a directed ligand-receptor term to standard gene-expression distances.
If this is right
- Improved cross-snapshot cell couplings compared with feature-only optimal transport.
- Better downstream trajectory estimates on both synthetic and real scRNA-seq datasets.
- Mechanistically interpretable in silico perturbations that alter trajectories by silencing ligand-receptor pairs.
- Trajectory shifts on lung cancer data that match expected effects of targeted pathway inhibition.
Where Pith is reading between the lines
- The same interaction-aware cost structure could be tested on snapshot data from other interacting systems, such as microbial communities or social populations.
- One could check whether the learned couplings predict outcomes of new perturbation experiments not used in training.
- Extending the interaction term to additional signaling modalities might further tighten the alignment without changing the overall transport framework.
Load-bearing premise
A directed interaction cost from ligand-receptor activity supplies biologically meaningful couplings beyond what gene-expression distances already capture.
What would settle it
No gain in coupling accuracy or trajectory quality on a dataset where cell communication is known to be irrelevant, or in silico perturbations that fail to match real pathway-inhibition outcomes, would falsify the central benefit.
Figures


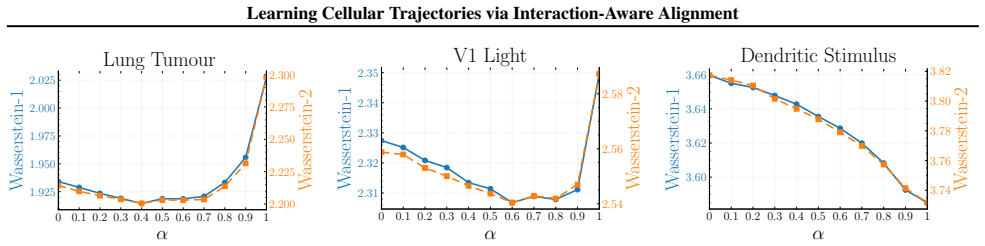



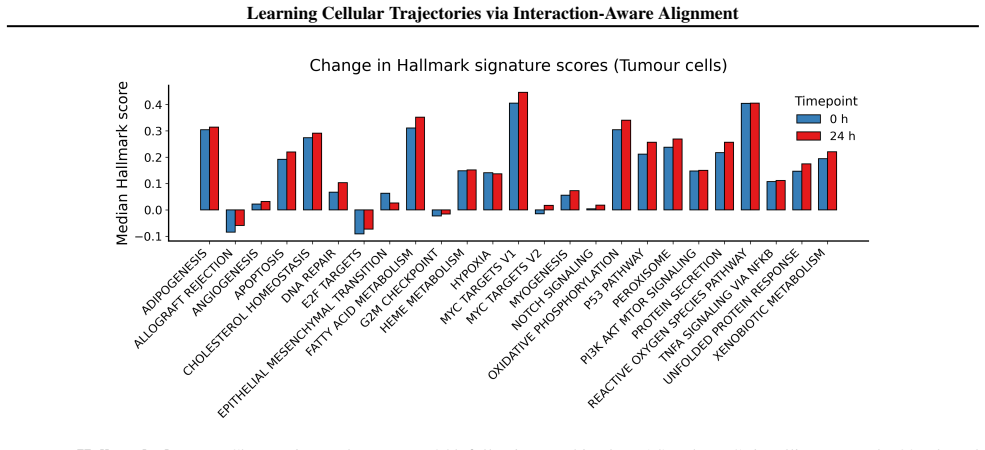
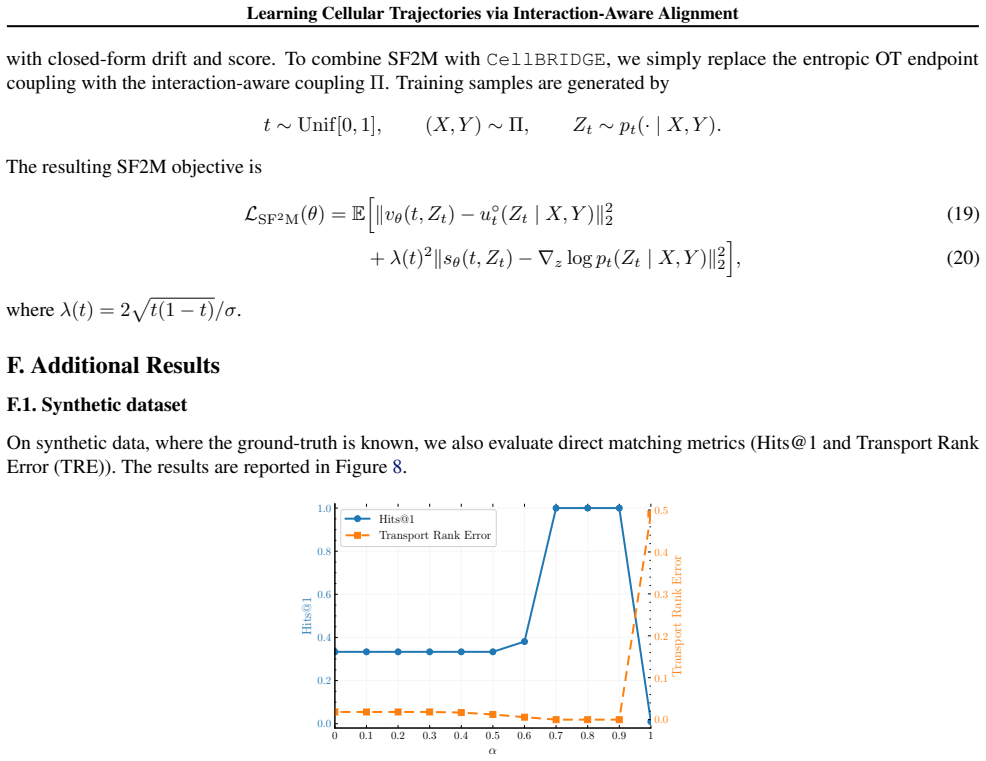
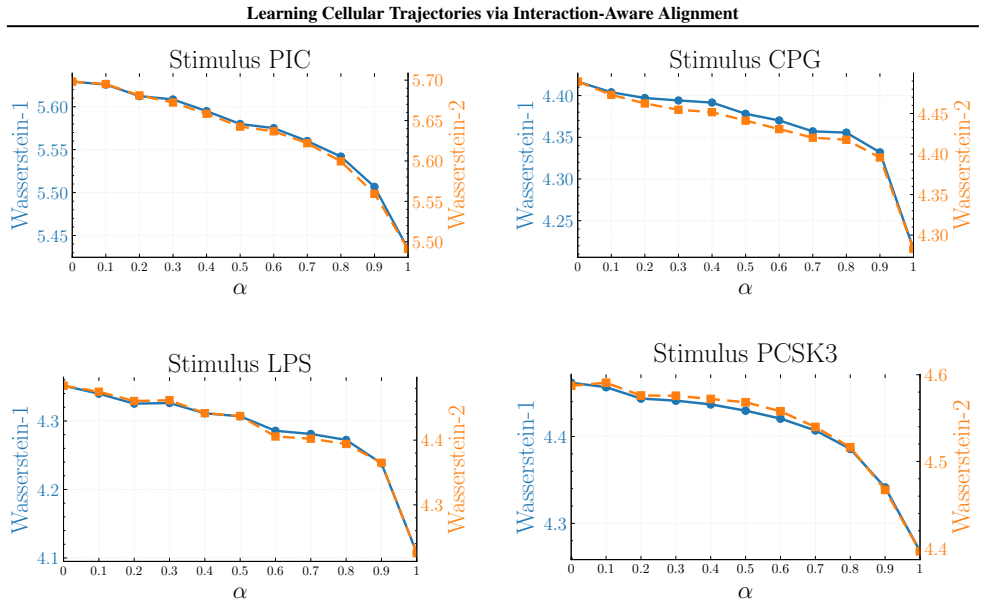
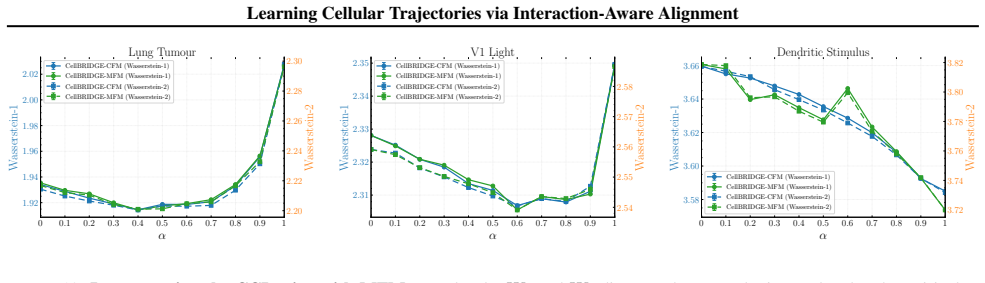
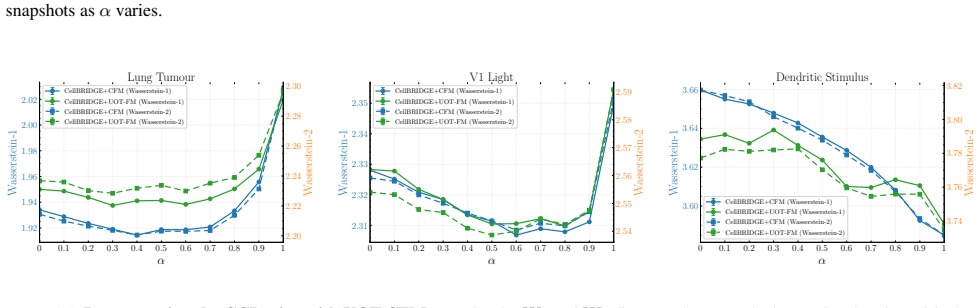
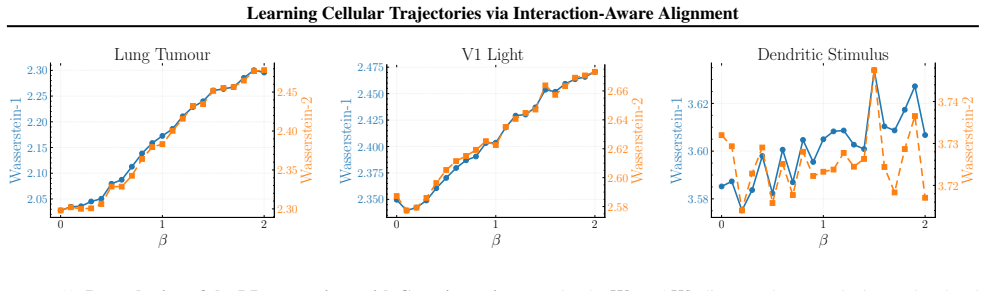
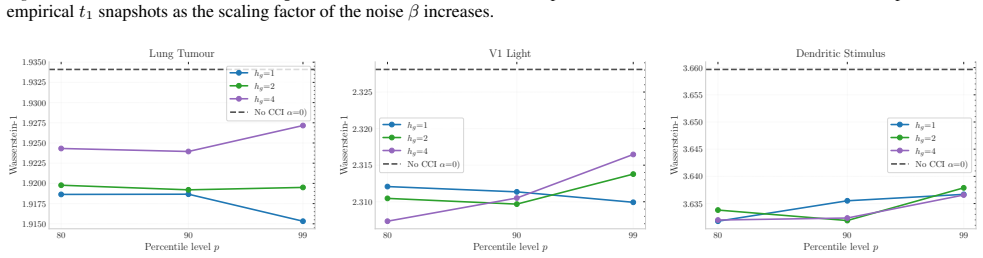
read the original abstract
Inferring dynamics from population snapshots is a fundamental challenge in machine learning and biology. In scRNA-sequencing (scRNA-seq), destructive measurements preclude direct tracking of individual cells across time, making trajectory inference underdetermined. Optimal Transport (OT) provides a principled framework for snapshot alignment, but a long-standing modeling question is which cost functions yield biologically meaningful couplings. Standard OT approaches rely on gene-expression distances, implicitly treating cells as independent points and neglecting structured cell-cell communication mediated by ligand-receptor signaling. We introduce CellBRIDGE (Cell-Based Regularized Interaction-Driven Gene Expression), which augments feature-based OT with a directed, typed interaction cost derived from ligand-receptor activity. By explicitly modeling cell-cell communication, CellBRIDGE improves cross-snapshot couplings and downstream trajectory estimates across synthetic and real scRNA-seq datasets relative to feature-only baselines. Notably, CellBRIDGE enables mechanistically interpretable in silico perturbations: on lung cancer data, silencing specific ligand-receptor pairs induces trajectory shifts that recapitulate expected effects of targeted pathway inhibition.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper introduces CellBRIDGE, which augments standard optimal transport for aligning scRNA-seq population snapshots by adding a directed, typed interaction cost derived from ligand-receptor activity to the feature-based cost. It claims this yields improved cross-snapshot couplings and trajectory estimates over expression-only baselines on synthetic and real datasets, plus the ability to perform mechanistically interpretable in silico perturbations (e.g., ligand-receptor silencing on lung cancer data that recapitulates expected pathway inhibition effects).
Significance. If the interaction term supplies couplings that are non-redundant with gene-expression distances, the method would provide a principled way to inject biological priors on cell-cell communication into trajectory inference, with the perturbation analysis offering a concrete route to mechanistic insight. The absence of any null-model control, however, leaves open whether reported gains reflect the claimed modeling of signaling or simply the effect of an additional regularizer.
major comments (2)
- [Abstract / Methods] The central claim that the ligand-receptor interaction cost produces biologically meaningful couplings beyond those already encoded in gene-expression distances is load-bearing, yet the manuscript contains no ablation that replaces the LR-derived matrix with a random matrix of matched sparsity and magnitude while keeping the OT solver fixed. Without this control, it is impossible to distinguish mechanistic contribution from generic regularization.
- [Abstract] No equations are supplied for the interaction cost (how ligand-receptor activity is quantified, normalized, or scaled relative to the expression kernel) or for the combined objective; this prevents verification that the two cost components are linearly independent or that the claimed improvements are not an artifact of the particular weighting chosen.
minor comments (2)
- The abstract and results sections should report error bars, data-split protocols, and statistical testing procedures for all quantitative claims.
- Notation for the interaction matrix and its integration into the transport plan should be introduced explicitly with a dedicated methods subsection.
Simulated Author's Rebuttal
We thank the referee for the constructive feedback highlighting the need for stronger controls and clearer exposition of the model. We address each major comment below and will revise the manuscript to incorporate the suggested additions.
read point-by-point responses
-
Referee: [Abstract / Methods] The central claim that the ligand-receptor interaction cost produces biologically meaningful couplings beyond those already encoded in gene-expression distances is load-bearing, yet the manuscript contains no ablation that replaces the LR-derived matrix with a random matrix of matched sparsity and magnitude while keeping the OT solver fixed. Without this control, it is impossible to distinguish mechanistic contribution from generic regularization.
Authors: We agree that this ablation is necessary to substantiate the claim that the gains arise from the biological structure of the LR interactions rather than from the addition of any structured regularizer. In the revised manuscript we will include an ablation that replaces the LR-derived cost matrix with a random matrix of identical sparsity and magnitude (while preserving the OT solver and all other hyperparameters) and report the resulting trajectory inference metrics on both the synthetic and real datasets. revision: yes
-
Referee: [Abstract] No equations are supplied for the interaction cost (how ligand-receptor activity is quantified, normalized, or scaled relative to the expression kernel) or for the combined objective; this prevents verification that the two cost components are linearly independent or that the claimed improvements are not an artifact of the particular weighting chosen.
Authors: The full mathematical definitions of the ligand-receptor activity quantification, the directed interaction cost matrix, its normalization, the scaling relative to the expression kernel, and the combined OT objective (including the weighting hyperparameter) appear in the Methods section. To address the concern about visibility and to facilitate verification of linear independence, we will move the key equations into the main text (or a dedicated Methods figure) in the revision. revision: yes
Circularity Check
No significant circularity in derivation chain
full rationale
The abstract and available text define CellBRIDGE as an augmentation of standard optimal transport using an interaction cost term sourced from external ligand-receptor databases rather than fitted or derived from the trajectory inference objective itself. No equations, self-citations, or ansatzes are presented that reduce any claimed prediction or result to the inputs by construction. The method is described as adding a directed typed cost to feature-based OT, with performance claims resting on empirical comparisons to baselines; this structure is self-contained against external benchmarks and does not invoke load-bearing self-citations or self-definitional steps.
Axiom & Free-Parameter Ledger
Reference graph
Works this paper leans on
-
[1]
ISSN 2662-8449. Camp, J. G., Badsha, F., Florio, M., Kanton, S., Gerber, T., Wilsch-Br¨auninger, M., Lewitus, E., Sykes, A., Hevers, W., Lancaster, M., Knoblich, J. A., Lachmann, R., P¨a¨abo, S., Huttner, W. B., and Treutlein, B. Human cerebral organoids recapitulate gene expression programs of fetal neocortex development.Proceedings of the National Acade...
-
[2]
ISSN 1091-6490. Cang, Z. and Nie, Q. Inferring spatial and signaling rela- tionships between cells from single-cell transcriptomic data.Nature Communications, 11(1):2084, 2020. Chen, G., Ren, C., Xiao, Y ., Wang, Y ., Yao, R., Wang, Q., You, G., Lu, M., Yan, S., Zhang, X., Zhang, J., Yao, Y ., and Zhou, H. Time-resolved single-cell transcriptomics reveals...
-
[3]
Fournier, N
URL https://github.com/PythonOT/ POT. Fournier, N. and Guillin, A. On the rate of convergence in Wasserstein distance of the empirical measure.Probabil- ity theory and related fields, 162(3):707–738, 2015. Goldfarbmuren, K. C., Jackson, N. D., Sajuthi, S. P., Dyjack, N., Li, K. S., Rios, C. L., Plender, E. G., Montgomery, M. T., Everman, J. L., Bratcher, ...
2015
-
[4]
Haghverdi, L., B ¨uttner, M., Wolf, F
ISSN 2041-1723. Haghverdi, L., B ¨uttner, M., Wolf, F. A., Buettner, F., and Theis, F. J. Diffusion pseudotime robustly reconstructs lineage branching.Nature Methods, 13(10):845–848, 2016. Hanahan, D. and Weinberg, R. A. Hallmarks of cancer: The next generation.Cell, 144(5):646–674, March 2011. ISSN 0092-8674. He, X. and Xu, C. Immune checkpoint signaling...
2041
-
[5]
ISSN 1748-7838. Hossain, I., Fanfani, V ., Fischer, J., Quackenbush, J., and Burkholz, R. Biologically informed NeuralODEs for genome-wide regulatory dynamics.Genome Biology, 25 (1):127, 2024. Hrvatin, S., Hochbaum, D. R., Nagy, M. A., Cicconet, M., Robertson, K., Cheadle, L., Zilionis, R., Ratner, A., Borges-Monroy, R., Klein, A. M., et al. Single-cell a...
work page internal anchor Pith review Pith/arXiv arXiv 2024
-
[6]
Wang, D., Jiang, Y ., Zhang, Z., Gu, X., Zhou, P., and Sun, J
Springer, 2008. Wang, D., Jiang, Y ., Zhang, Z., Gu, X., Zhou, P., and Sun, J. Joint Velocity-Growth Flow Matching for Single-Cell Dy- namics Modeling. InThe Thirty-ninth Annual Conference on Neural Information Processing Systems, 2025. Wang, S., Pisco, A. O., McGeever, A., Brbic, M., Zitnik, M., Darmanis, S., Leskovec, J., Karkanias, J., and Altman, R. B...
2008
-
[7]
Its complexity is O costOT(n0, n1) , (e.g
Linear OT subproblem.Given the linearized objective, we solve a linear OT problem over Π(a, b) using POT’s (Flamary et al., 2024) existing OT routine. Its complexity is O costOT(n0, n1) , (e.g. cubic innfor a network-simplex LP, orO(T Sinkhorn n0n1)for entropic OT). If TCG denotes the number of Frank–Wolfe iterations required to reach the desired toleranc...
2024
-
[8]
active” vs. “inactive
+n 0n1 , Hence it is quadratic in the number of cells per snapshot and linear in the number of LR pairs K. In comparison, a feature-only OT solver (α= 0) needsCandΓ, with memoryO(n 0n1). Using CellBRIDGE with large-scale datasets.While the computational and memory cost remained reasonable across the datasets we used, for very large datasets it can be miti...
2019
-
[9]
Analysis of the feature costF For the ground truth coupling ΓGT , each cluster k maps to its true image µ′ k. The cost is the mean squared norm of the translation vectorsv 0 = (4,−4),v 1 = (0,−4), andv 2 = (−4,−4): F(Γ GT ) = 1 3 2X k=0 ||vk||2 = 1 3(32 + 16 + 32) = 80 3 .(22) For the feature-only coupling ΓF O, S0 maps to S ′ 2, S1 to S ′ 1, and S2 to S ′
-
[10]
In the finite sample regime with N points, the optimal transport cost between two empirical Gaussian distributions with identical covariance matrices converges to the squared Euclidean distance between their means. We denote the finite-sample deviation byδ N : F(Γ F O) = 1 3 ||µ0 −µ ′ 2||2 +||µ 1 −µ ′ 1||2 +||µ 2 −µ ′ 0||2 +δ N .(23) Hence: F(Γ F O) = 16 ...
2015
-
[11]
Analysis of structure costS The structure cost is the Gromov-Wasserstein cost: S(Γ) = X i,k X j,l ||G(0) ik −G (1) jl ||2ΓijΓkl.(25) Since ΓGT maps every source cluster k to the target cluster with the same index k, and G(1) is defined to preserve the index-based structure ofG (0), we have: S(Γ GT ) = 0.(26) For ΓF O, the mapping permutes indices as π(0) ...
-
[12]
Threshold derivation with normalization We now incorporate the normalization scheme of Section D.4. Define the (unnormalized) feature and structure gaps between the two couplings as ∆F:=F(Γ F O)− F(Γ GT ),∆S:=S(Γ F O)− S(Γ GT ).(28) From the computations above, ∆F= 16 +δ N − 80 3 ,∆S= 4 9 .(29) For sufficiently large N, we have F(Γ GT )>F(Γ F O), so |∆F |...
-
[13]
The energyK(X ·)is minimized overA(Π Γ)by the process Xlin t := (1−t)X+tY,(X, Y)∼Π Γ.(42) 35 Learning Cellular Trajectories via Interaction-Aware Alignment
-
[14]
The minimal value of the kinetic energy is inf X·∈A(ΠΓ) K(X·) =E (X,Y)∼Π Γ ∥X−Y∥ 2 = X i,j Γij Cij.(43) Proof.AnyX · ∈ A(ΠΓ)satisfies(X 0, X1)∼Π Γ. Condition on the endpoints: K(X·) =E (X,Y)∼Π Γ " E hZ 1 0 ˙Xt 2 dt (X0, X1) = (X, Y) i# .(44) For each fixed pair (X, Y) = (x, y) , Lemma 1 shows that the conditional energy is minimized by the straight-line p...
-
[15]
For any fixedΓ, the inner infimum over X· ∈ A(ΠΓ) is attained by the straight-line process Xlin t = (1−t)X+tY , (X, Y)∼Π Γ, and inf X·∈A(ΠΓ) Eα(Γ, X·) = (1−α) X i,j ΓijCij +αS(Γ).(48)
-
[16]
Consequently, the joint static–dynamic problem reduces to the purely static FGW problem inf Γ∈Π(a,b) inf X·∈A(ΠΓ) Eα(Γ, X·) = inf Γ∈Π(a,b) h (1−α)⟨Γ, C⟩ F +αS(Γ) i ,(49) whose minimizers are exactly the FGW-optimal couplings used byCellBRIDGE. Proof.Point (1) follows directly from Proposition 1: for anyΓ, inf X·∈A(ΠΓ) Eα(Γ, X·) = (1−α) inf X·∈A(ΠΓ) K(X·) ...
2000
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.