Bilinear gating of motor primitives: a principle linking dendritic computation to rapid goal-directed adaptation
Pith reviewed 2026-06-27 10:48 UTC · model grok-4.3
The pith
In macaque motor cortex, neuron burst fraction encodes reach direction more selectively than firing rate because it computes the product of goal and state via dendritic coincidence detection.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
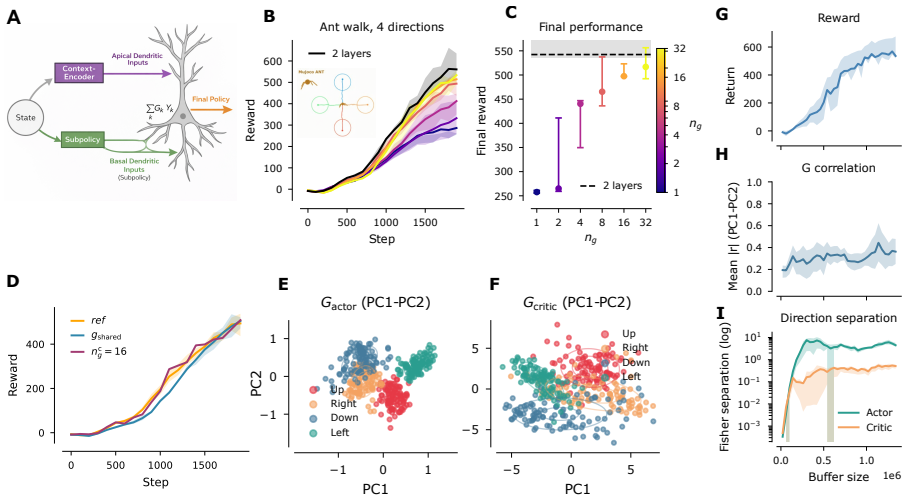
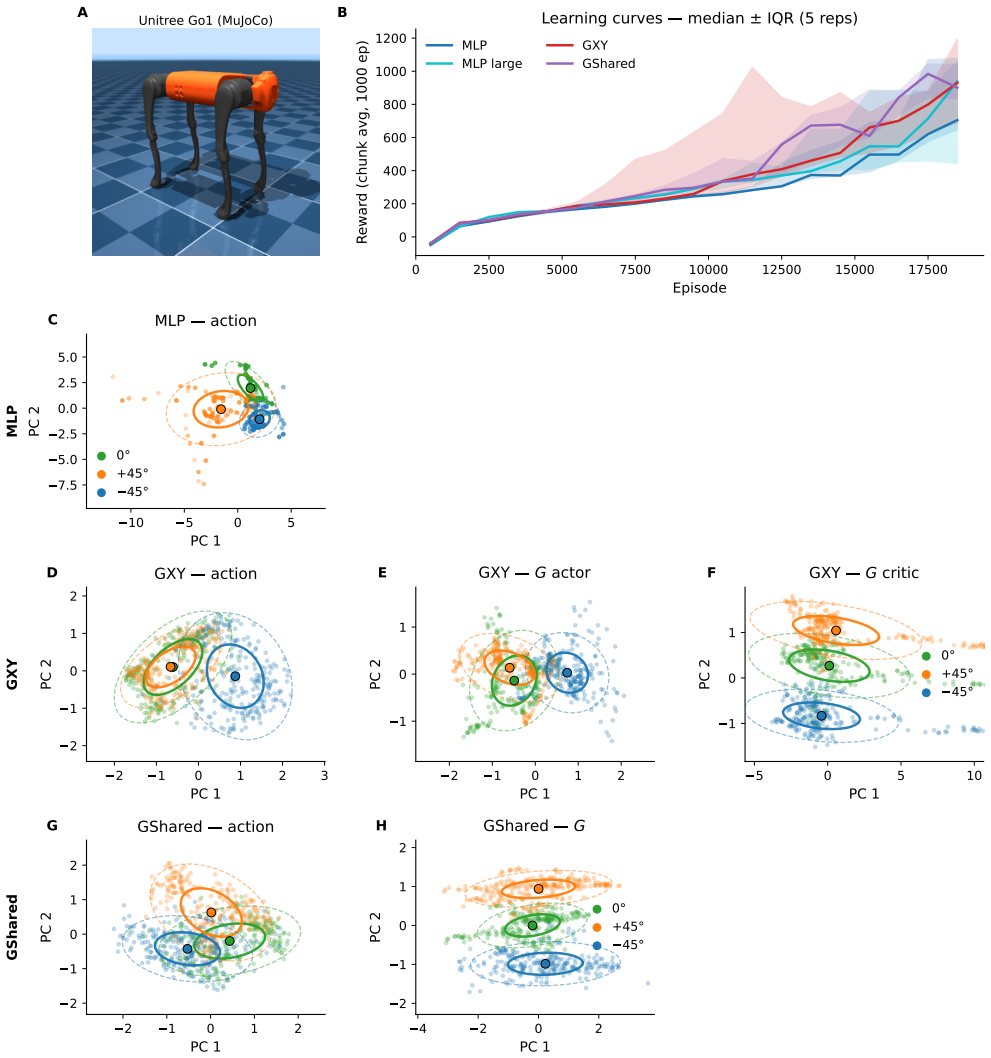
Burst fraction of neurons in motor cortex encodes reach direction far more selectively than firing rate, as a direct consequence of dendritic coincidence detection in layer-5 pyramidal neurons where goal-related apical input coincides with state-related basal drive, so that burst probability computes the bilinear gate G(g) Y(s). This same multiplicative gate, when used in a reinforcement-learning agent, supports zero-shot generalisation to new goals and rapid online adaptation.
What carries the argument
The bilinear gate G(g) Y(s) arising from coincidence of apical goal input and basal state drive in layer-5 pyramidal neurons, which gates bursting as the product of goal and state signals.
If this is right
- The observed dissociation between burst fraction and firing rate holds in every recording session across animals and labs.
- A minimal two-compartment spiking model reproduces the selective encoding in bursts.
- Embedding the multiplicative gate in an RL agent confers zero-shot generalisation to new goals.
- The mechanism provides a computational rationale for segregating goal information into bursts.
Where Pith is reading between the lines
- This form of gating could allow similar rapid adaptation in other cognitive tasks involving goal and state separation.
- Disrupting apical inputs specifically might eliminate the goal selectivity in bursts while preserving firing rates.
- Similar bilinear computations might be identifiable in other cortical regions using burst fraction analysis.
Load-bearing premise
The dissociation is produced by apical goal-related input coinciding with basal state-related drive in layer-5 pyramidal neurons rather than other cellular or network mechanisms.
What would settle it
Simultaneously controlling apical and basal inputs to layer-5 neurons in vitro and measuring whether burst probability equals the product of the two input strengths.
Figures





read the original abstract
Movement requires the motor cortex to specify both \emph{what} action to produce and \emph{which goal} it serves, yet how individual neurons separate these factors is not understood. Here we show that in macaque motor cortex the \emph{burst fraction} of a neuron, the proportion of its spikes emitted in high-frequency bursts, encodes reach direction far more selectively than its overall firing rate. This dissociation is highly consistent: it holds in every one of 12 recording sessions spanning three animals and two laboratories (all $p<10^{-12}$) and survives controls that remove any contribution of firing rate, showing that goal information is concentrated specifically in bursts. We then show that this coding signature is the predicted consequence of dendritic coincidence detection in layer-5 pyramidal neurons: when a goal-related apical input coincides with a state-related basal drive the neuron bursts, so burst probability computes the product of goal and state, a bilinear gate $G(g)\,Y(s)$. A minimal two-compartment spiking model reproduces the effect, and the same multiplicative gate, embedded in a reinforcement-learning agent, supports zero-shot generalisation to new goals and rapid online adaptation, providing a computational rationale for segregating goal information into bursts. These results identify burst fraction as a goal-selective code in motor cortex, tie it to a concrete cellular mechanism, and show that the mechanism confers a learning advantage.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript claims that in macaque motor cortex, the burst fraction of layer-5 pyramidal neurons encodes reach direction more selectively than mean firing rate (consistent across all 12 sessions in 3 animals with p<10^{-12} and surviving rate-matched controls), that this dissociation arises because burst probability implements a bilinear gate G(g)Y(s) via apical goal-related coincidence with basal state-related drive, that a minimal two-compartment spiking model reproduces the effect, and that embedding the same multiplicative gate in an RL agent yields zero-shot generalization to new goals and rapid online adaptation.
Significance. If the mechanistic interpretation is supported, the work supplies a concrete cellular-to-computational link between dendritic coincidence detection and goal-directed motor adaptation, with the statistical consistency across sessions and the RL demonstration of learning advantages constituting clear strengths. The result would be significant for theories that separate goal and state representations at the single-neuron level.
major comments (2)
- [Abstract / two-compartment spiking model] Abstract and the section presenting the two-compartment model: the central claim that burst probability computes the product G(g)Y(s) via apical-basal coincidence in L5 pyramids is an inference; the extracellular recordings establish only that burst fraction is more direction-selective than rate, while the model shows the dissociation is possible under the assumed compartmental assignment. No compartment-specific evidence (e.g., intracellular or optogenetic) is provided to rule out alternative cellular or circuit mechanisms (intrinsic bursting, inhibitory timing, or population synchrony), which is load-bearing for the proposed dendritic mechanism.
- [RL agent section] RL demonstration section: the claim that the multiplicative gate supports zero-shot generalization and rapid adaptation is presented as a computational rationale, yet the manuscript does not report whether the RL parameters were tuned to match the observed neural statistics or chosen independently; without this, it is unclear whether the performance advantage is a genuine prediction or a consequence of the modeling choices.
minor comments (2)
- [Methods] Methods: the controls that 'remove any contribution of firing rate' are described only at high level; explicit equations or pseudocode for how burst fraction is computed and how rate-matched surrogates are generated would improve clarity and reproducibility.
- [Figures] Figure legends: several panels compare burst fraction and rate selectivity but do not state the exact statistical test or correction used for the reported p<10^{-12} values across sessions.
Simulated Author's Rebuttal
We thank the referee for the constructive comments, which help sharpen the presentation of our claims. We respond to each major comment below.
read point-by-point responses
-
Referee: [Abstract / two-compartment spiking model] Abstract and the section presenting the two-compartment model: the central claim that burst probability computes the product G(g)Y(s) via apical-basal coincidence in L5 pyramids is an inference; the extracellular recordings establish only that burst fraction is more direction-selective than rate, while the model shows the dissociation is possible under the assumed compartmental assignment. No compartment-specific evidence (e.g., intracellular or optogenetic) is provided to rule out alternative cellular or circuit mechanisms (intrinsic bursting, inhibitory timing, or population synchrony), which is load-bearing for the proposed dendritic mechanism.
Authors: We agree that the bilinear-gate interpretation is a mechanistic hypothesis supported by the two-compartment model rather than a direct experimental demonstration from the extracellular data. The recordings establish the greater direction selectivity of burst fraction (with the reported statistical consistency), while the model shows that apical-basal coincidence can produce this dissociation. We will revise the abstract and model section to state explicitly that the dendritic mechanism is proposed on the basis of the model and is consistent with the data, while noting that alternative cellular or circuit mechanisms remain possible and are not ruled out by the present recordings. revision: yes
-
Referee: [RL agent section] RL demonstration section: the claim that the multiplicative gate supports zero-shot generalization and rapid adaptation is presented as a computational rationale, yet the manuscript does not report whether the RL parameters were tuned to match the observed neural statistics or chosen independently; without this, it is unclear whether the performance advantage is a genuine prediction or a consequence of the modeling choices.
Authors: The RL parameters were chosen independently of the recorded neural statistics to demonstrate the general computational benefit of the multiplicative gate; they were not fitted to the observed burst fractions or firing rates. We will add an explicit statement to this effect in the revised manuscript. revision: yes
Circularity Check
No significant circularity; derivation is deductive from independent observations
full rationale
The paper reports an empirical dissociation (burst fraction more direction-selective than mean rate) that is statistically robust across sessions and survives rate controls. It then deduces that apical-basal coincidence in L5 pyramids produces burst probability as the product G(g)Y(s), demonstrates this with a minimal two-compartment model constructed from standard dendritic properties, and shows that the same gate confers RL advantages. None of these steps reduce to a fit of the target data, a self-citation chain, or a renaming; the neural statistics are external to the model construction and the bilinear form follows directly from the coincidence logic without parameter tuning to the observed selectivity ratios.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Apical goal input coinciding with basal state drive produces high-frequency bursts in layer-5 pyramidal neurons.
Reference graph
Works this paper leans on
-
[1]
Thoroughman, K. A. & Shadmehr, R. Learning of action through adaptive combination of motor primitives.Nature407, 742–747 (2000)
2000
-
[2]
Larkum, M. E. A new cellular mechanism for coupling inputs to their outputs in pyramidal neurons.Nature Reviews Neuroscience14, 783–793 (2013)
2013
-
[3]
E., Zhu, J
Larkum, M. E., Zhu, J. J. & Sakmann, B. A new cellular mechanism for coupling inputs to their outputs in pyramidal neurons.Nature398, 338–341 (1999)
1999
-
[4]
& Segev, I
Hay, E., Hill, S., Schürmann, F., Markram, H. & Segev, I. Division of labor in the layer 5 apical dendrite.Frontiers in cellular neuroscience5, 28 (2011)
2011
-
[5]
Neuroscience research60, 268–279 (2008)
Urakubo, H.et al.Nonlinear dendritic integration in a functional model of rat layer 5 cortical pyramidal neuron. Neuroscience research60, 268–279 (2008)
2008
-
[6]
S., Anastassiou, C
Shai, A. S., Anastassiou, C. A., Larkum, M. E. & Koch, C. Physiology of layer 5 pyramidal neurons in mouse primary visual cortex: coincidence detection through bursting.PLoS Computational Biology11, e1004090 (2015). 8.Capone, C. & Falorsi, L. Adaptive behavior with stable synapses.Neural Networks108082 (2025)
2015
-
[7]
& Paolucci, P
Capone, C., Lupo, C., Muratore, P. & Paolucci, P. S. Beyond spiking networks: The computational advantages of dendritic amplification and input segregation.Proceedings of the National Academy of Sciences120, e2220743120 (2023). 10.Sherman, S. M. Thalamo-cortical interactions.Current Opinion in Neurobiology40, 78–84 (2016)
2023
-
[8]
& Markram, H
Maass, W., Natschäger, T. & Markram, H. Real-time computing without stable states: A new framework for neural computation based on perturbations.Neural computation14, 2531–2560 (2002)
2002
-
[9]
echo state
Jaeger, H. The "echo state" approach to analysing and training recurrent neural networks. Tech. Rep., German National Research Center for Information Technology (2001)
2001
-
[10]
& Strick, P
Rathelot, J.-A. & Strick, P. L. Motor cortical control of a fore-arm movement in 3d.Journal of Neuroscience29, 9859–9870 (2009)
2009
-
[11]
Rigoét, X.et al.The organization of prefrontal-subthalamic inputs in primates provides an anatomical substrate for both suppression and generation of action.Brain136, 1654–1667 (2013)
2013
-
[12]
& D’Esposito, M
Badre, D. & D’Esposito, M. Prefrontal cortex and hierarchical cognitive control.Annual review of neuroscience32, 167–191 (2009)
2009
-
[13]
Wise, S. P. The premotor cortex and the supplementary motor area: functional organisation.Trends in neurosciences8, 239–242 (1985)
1985
-
[14]
Sequential organization of multiple movements: involvement of cortical motor areas.Annual review of neuroscience 24, 631–651 (2001)
Tanji, J. Sequential organization of multiple movements: involvement of cortical motor areas.Annual review of neuroscience 24, 631–651 (2001). 18.Churchland, M. M.et al.Neural population dynamics during reaching.Nature487, 51–56 (2012)
2001
-
[15]
InNeurIPS 2021 Datasets and Benchmarks Track(2021)
Pei, F.et al.Neural latents benchmark ’21: Evaluating latent variable models of neural population activity. InNeurIPS 2021 Datasets and Benchmarks Track(2021)
2021
-
[16]
& Sprekeler, H
Naud, R. & Sprekeler, H. Sparse bursts optimize information transmission in a multiplexed neural code.Proceedings of the National Academy of Sciences115, E6329–E6338 (2018)
2018
-
[17]
Payeur, A., Guerguiev, J., Zenke, F., Richards, B. A. & Naud, R. Burst-dependent synaptic plasticity can coordinate learning in hierarchical circuits.Nature Neuroscience24, 1010–1019 (2021)
2021
-
[18]
G., Hegemann, P
Takahashi, N., Oertner, T. G., Hegemann, P. & Larkum, M. E. Active cortical dendrites modulate perception.Science354, 1587–1590 (2016)
2016
-
[19]
W., Pemberton, J., Mellor, J
Greedy, W., Zhu, H. W., Pemberton, J., Mellor, J. & Costa, R. P. Single-phase deep learning in cortico-cortical networks. InAdvances in Neural Information Processing Systems, vol. 35 (2022)
2022
-
[20]
Probabilistic logics and the synthesis of reliable organisms from unreliable components
von Neumann, J. Probabilistic logics and the synthesis of reliable organisms from unreliable components. In Shannon, C. E. & McCarthy, J. (eds.)Automata Studies, 43–98 (Princeton University Press, Princeton, NJ, 1956)
1956
-
[21]
Gaines, B. R. Stochastic computing systems. In Tou, J. T. (ed.)Advances in Information Systems Science, vol. 2, 37–172 (Plenum Press, New York, 1969)
1969
-
[22]
InAdvances in Neural Information Processing Systems (NeurIPS)(2017)
Barreto, A.et al.Successor features for transfer in reinforcement learning. InAdvances in Neural Information Processing Systems (NeurIPS)(2017)
2017
-
[23]
Barreto, A.et al.Transfer in reinforcement learning with successor features and generalized policy improvement.Journal of Machine Learning Research19, 1–64 (2018)
2018
-
[24]
M., Diedrichsen, J
Wolpert, D. M., Diedrichsen, J. & Flanagan, J. R. Principles of sensorimotor learning.Nature Reviews Neuroscience12, 739–751 (2011)
2011
-
[25]
Todorov, E., Erez, T. & Tassa, Y . MuJoCo: A physics engine for model-based control. In2012 IEEE/RSJ International Conference on Intelligent Robots and Systems (IROS), 5026–5033 (IEEE, 2012). 18/19 30.Brockman, G.et al.OpenAI gym (2016).1606.01540
work page internal anchor Pith review Pith/arXiv arXiv 2012
-
[26]
& Levine, S
Haarnoja, T., Zhou, A., Abbeel, P. & Levine, S. Soft actor-critic: Off-policy maximum entropy deep reinforcement learning with a stochastic actor. InProceedings of the 35th International Conference on Machine Learning (ICML), 1861–1870 (PMLR, 2018). 19/19
2018
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.