Begging with a Purpose? Testing Behavioural Hallmarks of First-Order Intentionality in Free-ranging Hanuman Langurs
Pith reviewed 2026-06-27 05:08 UTC · model grok-4.3
The pith
Free-ranging Hanuman langurs display multiple behavioral hallmarks of first-order intentionality in their human-directed gestures.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
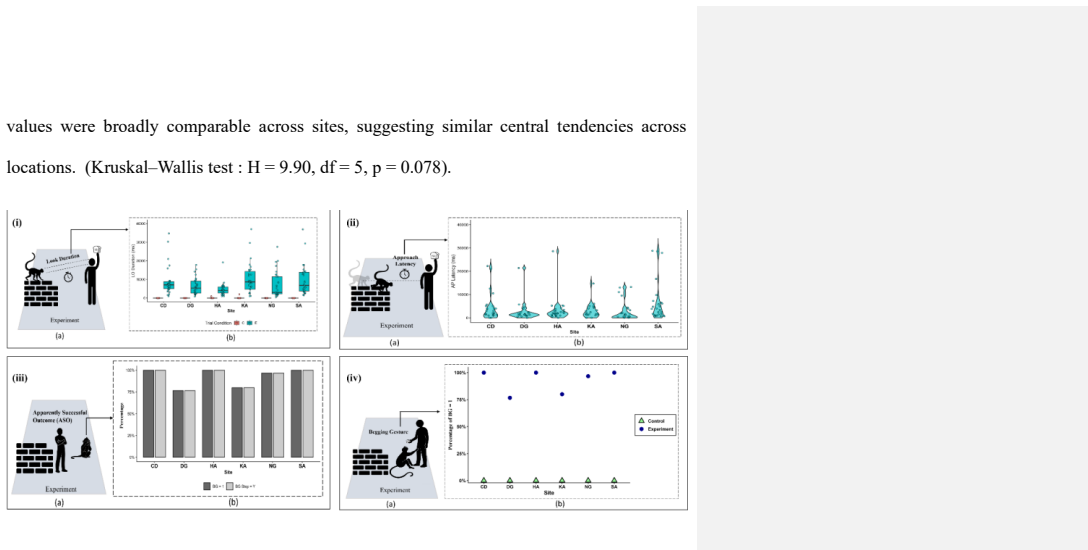
Experimental trials elicited audience checking, recipient-directed orientation, rapid approach responses, food-solicitation gestures and gestural flexibility, whereas these behaviours were rare or absent in control trials. Differences between experimental and control conditions were significant across all six study sites. Signalling also ceased following food acquisition, consistent with the stopping rule associated with an Apparently Satisfactory Outcome. Our findings demonstrate the presence of multiple behavioural hallmarks linked to first-order intentionality in the human-directed gestural communication of free-ranging Hanuman langurs.
What carries the argument
The set of behavioural markers used to operationalize first-order intentionality: audience checking, recipient-directed orientation, rapid approach responses, food-solicitation gestures, gestural flexibility, and the stopping rule after an Apparently Satisfactory Outcome.
If this is right
- Intentional communication occurs in free-ranging non-ape primates.
- The evolutionary distribution of intentionality-related traits extends across primates beyond apes.
- Human-directed gestures in wild langurs can be tested experimentally for cognitive markers.
- The observed patterns remain consistent across multiple anthropogenic sites.
Where Pith is reading between the lines
- Similar experiments could test for these markers in other monkey species that interact with humans.
- The results suggest that proximity to humans may influence how primates develop intentional signaling.
- Future comparisons between free-ranging and captive groups could clarify environmental effects on these behaviors.
Load-bearing premise
The quantified markers validly capture first-order intentionality rather than reflexive or learned responses without mental-state attribution.
What would settle it
If audience checking, flexible gesturing, and the stopping rule after food acquisition appeared at similar rates in control trials without food presentation.
Figures



read the original abstract
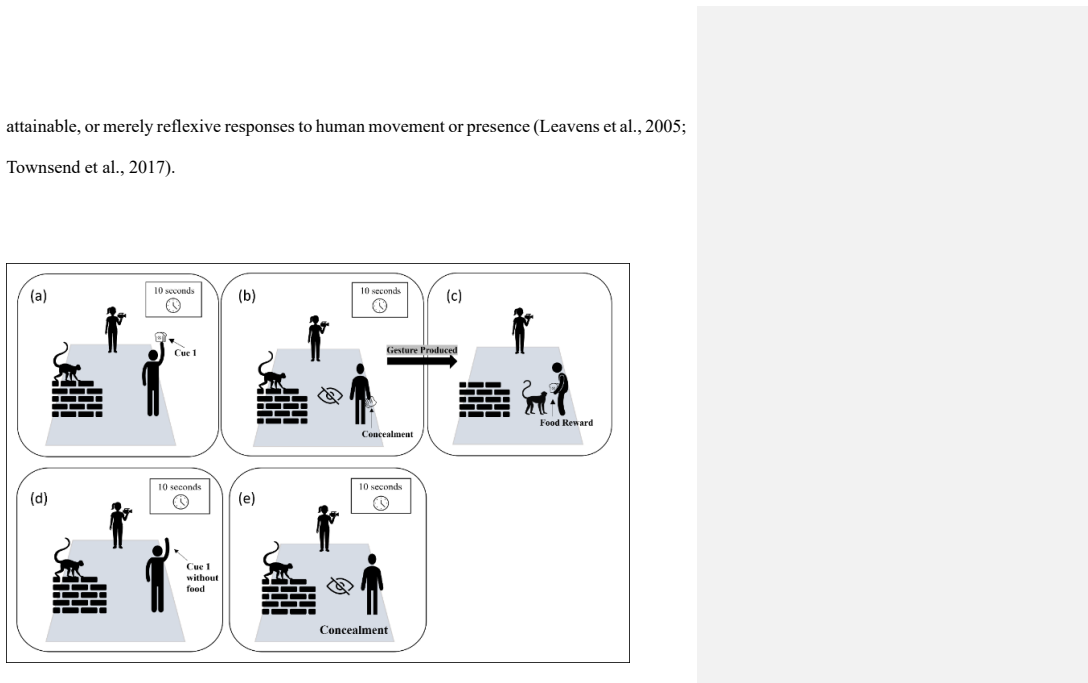
Intentional communication has been studied extensively in primates, yet evidence from free-ranging non-ape species remains limited. Human-directed food-solicitation gestures in Hanuman langurs (Semnopithecus entellus) have recently been described, but whether these behaviours exhibit behavioural hallmarks associated with first-order intentionality remains unknown. Here, we experimentally investigated the presence of these hallmarks in free-ranging Hanuman langurs across six anthropogenic sites in southern West Bengal, India. We conducted 360 experimental and control trials and quantified behavioural markers commonly used to operationalize intentional communication. Experimental trials elicited audience checking, recipient-directed orientation, rapid approach responses, food-solicitation gestures and gestural flexibility, whereas these behaviours were rare or absent in control trials. Differences between experimental and control conditions were significant across all six study sites. Signalling also ceased following food acquisition, consistent with the stopping rule associated with an Apparently Satisfactory Outcome. Our findings demonstrate the presence of multiple behavioural hallmarks linked to first-order intentionality in the human-directed gestural communication of free-ranging Hanuman langurs. These results extend the study of intentionality beyond apes and provide new insights into the evolutionary distribution of intentionality-related traits across primates.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript reports results from 360 experimental and control trials across six anthropogenic sites in India testing whether human-directed food-solicitation gestures by free-ranging Hanuman langurs exhibit behavioral hallmarks of first-order intentionality. Markers quantified include audience checking, recipient-directed orientation, rapid approach responses, food-solicitation gestures, gestural flexibility, and cessation of signaling after an Apparently Satisfactory Outcome (food acquisition). The authors report these behaviors occur significantly more often in experimental (food-present) than control trials at all sites and conclude that the data demonstrate first-order intentionality in a non-ape primate species.
Significance. If the central interpretation holds, the multi-site empirical design would extend the documented distribution of intentional-communication markers beyond apes to colobines and strengthen comparative claims about the evolutionary history of these traits. The use of free-ranging subjects and a stopping-rule test are positive features of the study.
major comments (2)
- [Methods / Experimental Design] The experimental contrast (food-present vs. food-absent) does not include controls capable of dissociating first-order intentional signaling from associative learning or reflexive orientation to salient human cues (e.g., non-social food cues, habituation trials, or non-communicative goal-directed actions). Because the same behavioral sequence is predicted by simpler reinforcement mechanisms at anthropogenic sites, this omission is load-bearing for the claim that the quantified markers specifically operationalize first-order intentionality.
- [Abstract / Results] The abstract states that differences were significant across all six sites and consistent with the stopping rule, yet reports no statistical tests, effect sizes, inter-observer reliability coefficients, or controls for site or individual identity. These details are required to evaluate whether the data support the cross-site consistency claim.
minor comments (1)
- [Methods] The operational definitions of each behavioral marker (audience checking, gestural flexibility, etc.) should be stated explicitly with reference to prior literature so that readers can assess their validity independent of the intentionality interpretation.
Simulated Author's Rebuttal
We thank the referee for their detailed and constructive comments on our manuscript. We address each major point below and outline planned revisions where appropriate.
read point-by-point responses
-
Referee: [Methods / Experimental Design] The experimental contrast (food-present vs. food-absent) does not include controls capable of dissociating first-order intentional signaling from associative learning or reflexive orientation to salient human cues (e.g., non-social food cues, habituation trials, or non-communicative goal-directed actions). Because the same behavioral sequence is predicted by simpler reinforcement mechanisms at anthropogenic sites, this omission is load-bearing for the claim that the quantified markers specifically operationalize first-order intentionality.
Authors: We agree that additional controls could further isolate intentionality from associative or reflexive processes. However, our design follows standard protocols used in ape intentionality studies (e.g., audience checking and stopping-rule tests), and the food-absent condition controls for baseline human-directed behaviors at the same sites. The observed patterns—particularly gestural flexibility, recipient-directed orientation, and cessation after food acquisition—are less parsimoniously explained by simple reinforcement alone. We will add an explicit discussion of this limitation and its implications for interpretation in the revised manuscript. revision: partial
-
Referee: [Abstract / Results] The abstract states that differences were significant across all six sites and consistent with the stopping rule, yet reports no statistical tests, effect sizes, inter-observer reliability coefficients, or controls for site or individual identity. These details are required to evaluate whether the data support the cross-site consistency claim.
Authors: The full manuscript reports the relevant statistical tests (including site-level comparisons), effect sizes, inter-observer reliability (Cohen’s kappa), and mixed-effects models controlling for site and individual identity in the Results and Statistical Analysis sections. The abstract is a concise summary and does not include these values due to length constraints. To improve clarity, we will revise the abstract to reference the key statistical support for cross-site consistency while retaining the word limit. revision: yes
Circularity Check
No circularity: empirical comparison of pre-defined markers across conditions
full rationale
The paper reports an experimental study that applies established behavioral markers (audience checking, recipient-directed orientation, etc.) drawn from prior primate communication literature to new data from controlled food-present vs. food-absent trials. These markers are treated as operational definitions imported from external sources rather than derived or fitted within the paper itself. No equations, parameter estimation, or self-citation chains are used to generate the central claim; the result is a straightforward frequency comparison that stands or falls on the external validity of the imported markers. No load-bearing step reduces to a self-definition or fitted input by construction.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Behavioral markers such as audience checking, recipient-directed orientation, gestural flexibility, and the stopping rule after food acquisition operationalize first-order intentionality.
Reference graph
Works this paper leans on
-
[1]
Altmann, J. (1974). Observational study of behavior: sampling methods. Behaviour, 49(3‑4), 227–266
1974
-
[2]
R., Kuroshima, H., Hattori, Y ., & Fujita, K
Anderson, J. R., Kuroshima, H., Hattori, Y ., & Fujita, K. (2010). Flexibility in the use of requesting gestures in squirrel monkeys (Saimiri sciureus). American Journal of Primatology, 72(8), 707–714. https://doi.org/10.1002/ajp.20827
-
[3]
A., Liebal, K., & Pika, S
Arbib, M. A., Liebal, K., & Pika, S. (2008). Primate vocalization, gesture, and the evolution of human language. Current Anthropology, 49(6), 1053–1076
2008
-
[4]
Bard, K. A. (1992). Intentional behavior and intentional communication in young free‑ranging orangutans. Child Development, 63(5), 1186 –1197. https://doi.org/10.1111/j.1467-8624.1992.tb01688.x
-
[5]
Bates, E., Camaioni, L., & V olterra, V . (1975). The acquisition of performatives prior to speech. Merrill‑Palmer Quarterly of Behavior and Development, 21 (3), 205–226
1975
-
[6]
Ben Mocha, Y ., & Burkart, J. M. (2021). Intentional communication: Solving methodological issues to assigning first‑order intentional signalling. Biological Reviews, 96(3), 903–921. https://doi.org/10.1111/brv.12685
-
[7]
Bourjade, M., Meguerditchian, A., Maille, A., Gaunet, F., & Vauclair, J. (2014). Olive baboons, Papio anubis, adjust their visual and auditory intentional gestures to the visual 25 attention of others. Animal Behaviour, 87, 121 –128. https://doi.org/10.1016/j.anbehav.2013.10.019
-
[8]
Brentano, F. (1874). Psychologie vom empirischen Standpunkt. Duncker & Humblot. https://doi.org/10.1515/9783110332582
-
[9]
Bruner, J. S. (1981). Intention in the structure of action and interaction. Advances in Infancy Research, 1, 41–56
1981
-
[10]
W., Cartmill, E., Genty, E., Graham, K
Byrne, R. W., Cartmill, E., Genty, E., Graham, K. E., Hobaiter, C., & Tanner, J. (2017). Great ape gestures: Intentional communication with a rich set of innate signals. Animal Cognition, 20(4), 755–769. https://doi.org/10.1007/s10071-017-1096-4
-
[11]
Call, J., & Tomasello, M. (Eds.). (2007). The gestural communication of apes and monkeys. Lawrence Erlbaum Associates
2007
-
[12]
Canteloup, C., Bovet, D., & Meunier, H. (2015). Intentional gestural communication and discrimination of human attentional states in rhesus macaques (Macaca mulatta). Animal Cognition, 18(6), 875–883. https://doi.org/10.1007/s10071-015-0856-2
-
[13]
A., & Byrne, R
Cartmill, E. A., & Byrne, R. W. (2007). Orangutans modify their gestural signaling according to their audience’s comprehension. Current Biology, 17(15), 1345–1348
2007
-
[14]
A., & Byrne, R
Cartmill, E. A., & Byrne, R. W. (2010). Semantics of primate gestures: intentional meanings of orangutan gestures. Animal Cognition, 13(6), 793–804
2010
-
[15]
Cohen, J. (1960). A coefficient of agreement for nominal scales. Educational and psychological measurement, 20(1), 37-46
1960
-
[16]
Dasgupta, D., Banerjee, Ar., Dutta, A., Mitra, S., Banerjee, De., Karar, R., Karmakar, S., Bhattacharya, A., Ghosh, S., Bhattacharjee, P., & Paul, M. (2025). Decoding food solicitation techniques applied by free‑ranging Hanuman langurs residing in an urb an habitat. Animal Cognition, 28, 25. https://doi.org/10.1007/s10071-024-01925-y 26
-
[17]
Dennett, D. C. (1983). Intentional systems in cognitive ethology: The “Panglossianparadigm” defended. Behavioral and Brain Sciences, 6, 343 –390. https://doi.org/10.1017/S0140525X00016393
-
[18]
Deshpande, A., Gupta, S., & Sinha, A. (2018). Intentional communication between wild bonnet macaques and humans. Scientific Reports, 8 (1), 5147. https://doi.org/10.1038/s41598-018-22928-z
-
[19]
Genty, E., Breuer, T., Hobaiter, C., & Byrne, R. W. (2009). Gestural communication of the gorilla (Gorilla gorilla): Repertoire, intentionality and possible origins. Animal Cognition, 12(3), 527–546. https://doi.org/10.1007/s10071-009-0213-4
-
[20]
Graham, K. E., Furuichi, T., & Byrne, R. W. (2017). The gestural repertoire of the wild bonobo (Pan paniscus): A mutually understood communication system. Animal Cognition, 20(2), 171–177. https://doi.org/10.1007/s10071-016-1035-9
-
[21]
Graham, K. E., Wilke, C., Lahiff, N. J., & Slocombe, K. E. (2019). Scratching beneath the surface: Intentionality in great ape signal production. Philosophical Transactions of the Royal Society B: Biological Sciences, 375, 20180403. https://doi.org/10.1098/rstb.2018.0403
-
[22]
Grice, H. P. (1957). Meaning. The Philosophical Review, 66, 377–388. https://doi.org/10.2307/2182440
-
[23]
Gupta, S., & Sinha, A. (2019). Gestural communication of wild bonnet macaques in the Bandipur National Park, Southern India. Behavioural Processes, 168, 103956. https://doi.org/10.1016/j.beproc.2019.103956
-
[24]
Hobaiter, C., & Byrne, R. W. (2014). The meanings of chimpanzee gestures. Current Biology, 24(14), 1596–1600. https://doi.org/10.1016/j.cub.2014.05.066 27
-
[25]
Leavens, D. A., Bard, K. A., & Hopkins, W. D. (2019). The mismeasure of ape social cognition. Animal Cognition, 22 (4), 487 –504. https://doi.org/10.1007/s10071-017- 1119-1
-
[26]
Leavens, D. A., & Hopkins, W. D. (1998). Intentional communication by chimpanzees: A cross‑sectional study of the use of referential gestures. Developmental Psychology, 34 (5), 813–822. https://doi.org/10.1037/0012-1649.34.5.813
-
[27]
Leavens, D. A., Hostetter, A. B., Wesley, M. J., & Hopkins, W. D. (2004). Tactical use of unimodal and bimodal communication by chimpanzees (Pan troglodytes). Animal Behaviour, 67 (3), 467–476. https://doi.org/10.1016/j.anbehav.2003.04.007
-
[28]
Liebal, K., Waller, B. M., Burrows, A. M., & Slocombe, K. E. (2013). Primate communication: A multimodal approach. Cambridge University Press. https://doi.org/10.1016/j.applanim.2016.02.005
-
[29]
Maille, A., Engelhart, L., Bourjade, M., & Blois‑Heulin, C. (2012). To beg, or not to beg? That is the question: Mangabeys modify their production of requesting gestures in response to human’s attentional states. PLoS ONE, 7 (7), e41197.https://doi.org/10.1371/journal.pone.0041197
-
[30]
Meunier, H., Prieur, J., & Vauclair, J. (2013). Olive baboons communicate intentionally by pointing. Animal Cognition, 16 (2), 155 –163. https://doi.org/10.1007/s10071-012- 0558-y
-
[31]
Molesti, S., Meguerditchian, A., & Bourjade, M. (2020). Gestural communication in olive baboons (Papio anubis): Repertoire and intentionality. Animal Cognition, 23 (1), 19–40. https://doi.org/10.1007/s10071-019-01312-y
-
[32]
Rodrigues, E. D., & Fröhlich, M. (2024). Operationalizing intentionality in primate communication: Social and ecological considerations. International Journal of Primatology, 45, 501–525. https://doi.org/10.1007/s10764-021-00248-w 28
-
[33]
Rodrigues, E. D., Santos, A. J., Veppo, F., Pereira, J., & Hobaiter, C. (2021). Connecting primate gesture to the evolutionary roots of language: A systematic review. American Journal of Primatology, 83 (9), e23313. https://doi.org/10.1002/ajp.23313
-
[34]
M., Bono, A., Aychet, J., Pika, S., & Lemasson, A
Schel, A. M., Bono, A., Aychet, J., Pika, S., & Lemasson, A. (2022). Intentional gestural communication amongst red‑capped mangabeys (Cercocebus torquatus). Animal Cognition, 25, 1313–1330. https://doi.org/10.1007/s10071-022-01615-7
-
[35]
Scott‑Phillips, T. C. (2015). Meaning in animal and human communication. Animal Cognition, 18, 801–805. https://doi.org/10.1007/s10071-015-0845-5
-
[36]
Tomasello, M. (2008). Origins of human communication. MIT Press. https://doi.org/10.1111/j.1468-0017.2009.01388.x
-
[37]
Townsend, S. W., et al. (2017). Exorcising Grice’s ghost: An empirical approach to studying intentional communication in animals. Biological Reviews, 92, 1427 –1433. https://doi.org/10.1111/brv.12289
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.