Oscillations and Spatial Patterns in Large-Scale Stochastic Gene Regulatory Networks
Pith reviewed 2026-06-26 15:09 UTC · model grok-4.3
The pith
Fluctuations can induce stochastic Turing instability in gene networks that remain stable without noise.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
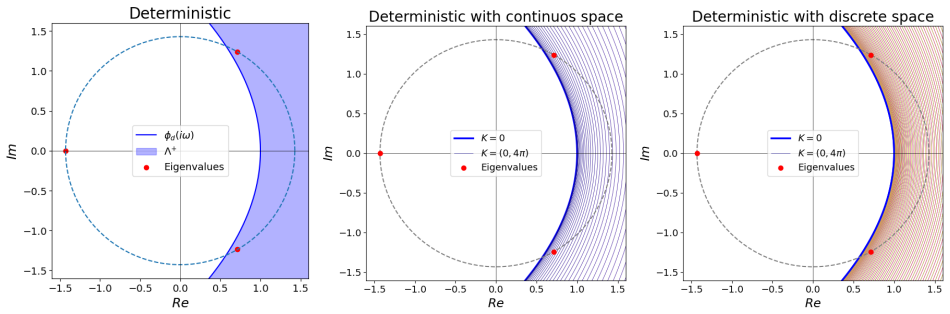
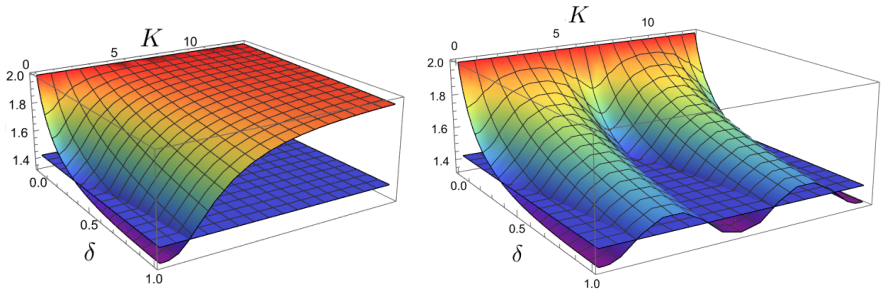
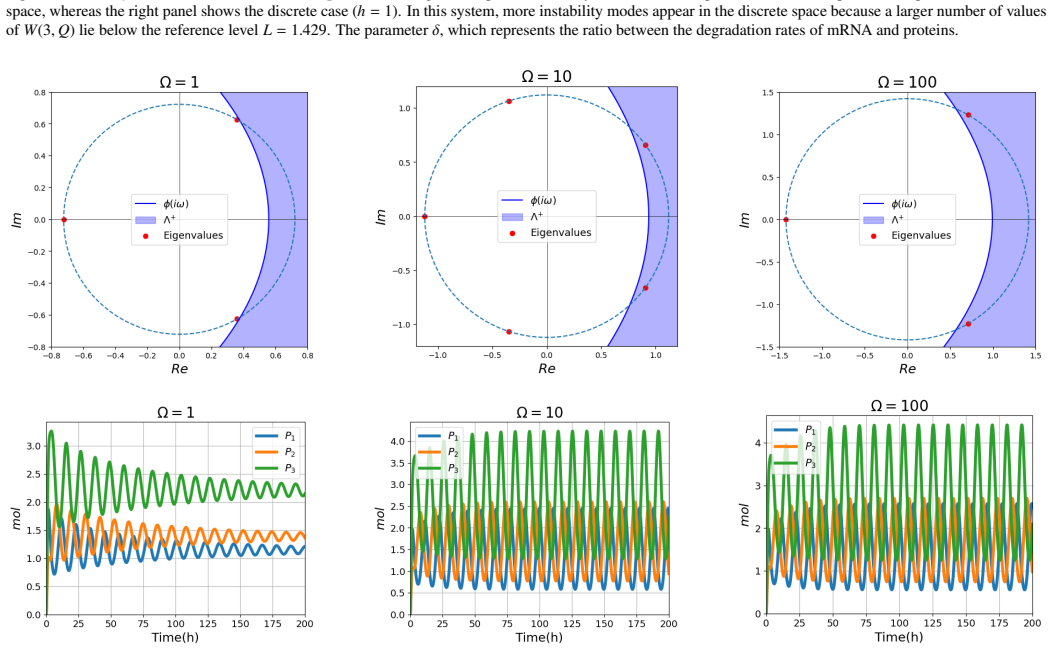
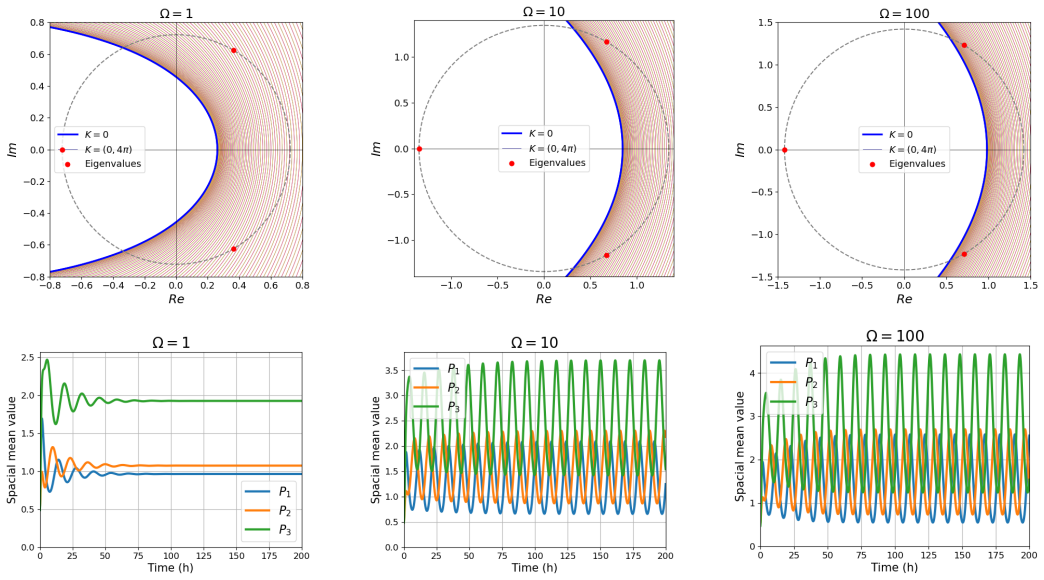
The stochastic framework based on the second-moment approach reveals that for small system sizes, fluctuations can dominate the dynamics and induce stochastic Turing instability, even when the system is stable in the absence of diffusion. Notably, Turing instabilities can emerge even when all variables have the same diffusion rate. The developed framework provides a systematic method for analyzing the stability of high-dimensional stochastic systems with diffusion, thereby simplifying the prediction of Turing and Turing-Hopf instabilities.
What carries the argument
Second-moment closure approximation applied to the stochastic dynamics of cyclic negative-feedback GRNs with diffusion.
Load-bearing premise
The second-moment closure accurately describes the stochastic dynamics and the network is strictly cyclic with negative feedback.
What would settle it
Direct observation that a small cyclic negative-feedback gene circuit with equal diffusion coefficients for all species remains spatially uniform when its deterministic version is stable would falsify the instability prediction.
Figures





read the original abstract
Gene regulatory networks (GRNs) are fundamental to cellular growth and tissue formation, orchestrating spatially and temporally regulated gene expression during development. These networks are inherently subject to intrinsic fluctuations arising from molecular noise, making the analysis of their stability essential for understanding robust pattern formation and developmental dynamics of the organism. In this study, we analyze the stability and dynamics of cyclic GRNs with negative feedback and diffusion, considering both deterministic and stochastic approaches. In the deterministic case, the system exhibits a bifurcation between stability and instability, leading to Hopf instability in the absence of diffusion and to Turing-Hopf instability when diffusion is included. It was observed that the discretization of the spatial domain introduces additional unstable modes, enabling a wider range of patterns. The stochastic framework based on the second-moment approach, which incorporates intrinsic fluctuations, reveals that for small system sizes, fluctuations can dominate the dynamics and induce stochastic Turing instability, even when the system is stable in the absence of diffusion. Notably, Turing instabilities can emerge even when all variables have the same diffusion rate. The developed framework provides a systematic method for analyzing the stability of high-dimensional stochastic systems with diffusion, thereby simplifying the prediction of Turing and Turing-Hopf instabilities. These findings contribute to a deeper understanding of the complex dynamics and pattern formation in GRNs, with potential implications for biological processes, such as cellular differentiation and development.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript analyzes cyclic gene regulatory networks with negative feedback, both deterministically (showing Hopf instability without diffusion and Turing-Hopf with diffusion, plus effects of spatial discretization) and stochastically via a second-moment closure on the chemical master equation with diffusion. The central claim is that intrinsic fluctuations induce stochastic Turing instability for small system sizes, even when the deterministic system is stable and even when all species have identical diffusion coefficients; the framework is presented as simplifying stability analysis for high-dimensional stochastic reaction-diffusion systems.
Significance. If the second-moment closure remains accurate in the fluctuation-dominated regime, the work supplies a systematic, computationally tractable route to predicting stochastic Turing and Turing-Hopf instabilities in large cyclic GRNs, extending classical deterministic pattern-formation theory to intrinsically noisy molecular networks with potential relevance to developmental patterning.
major comments (3)
- [§3.2] §3.2 (second-moment closure): the truncation after the covariance equations is used to derive the effective linear stability matrix whose eigenvalues determine the stochastic Turing threshold, yet no a-priori bound or numerical check is supplied showing that third- and higher-order moments remain negligible precisely when system size N is small enough for fluctuations to dominate; the eigenvalue crossing that produces the claimed instability therefore inherits an uncontrolled approximation error.
- [§4.1] §4.1, Eq. (18)–(22): the stochastic dispersion relation is obtained by closing the moment hierarchy and then taking the continuum limit; because the closure error scales with 1/N and the instability is asserted to appear at small N, the deterministic limit (N→∞) and the fluctuation-induced instability cannot be simultaneously valid without an explicit error estimate or comparison to exact stochastic simulations (e.g., Gillespie SSA on the same lattice).
- [Table 1, Fig. 3] Table 1 and Fig. 3: the reported parameter sets that produce equal-diffusion stochastic Turing patterns are obtained under the closed second-moment dynamics; without an independent verification that the same parameters produce spatial patterns in the full stochastic model, the claim that “Turing instabilities can emerge even when all variables have the same diffusion rate” rests on the untested closure.
minor comments (2)
- [Abstract] The abstract states that “the discretization of the spatial domain introduces additional unstable modes” but the corresponding linear-algebra argument is only sketched; a short appendix deriving the discrete Laplacian eigenvalues would clarify the claim.
- [§2.1] Notation for the diffusion matrix D and the Jacobian J is introduced without an explicit statement of their dimensions or the ordering of species; adding a sentence in §2.1 would remove ambiguity for readers.
Simulated Author's Rebuttal
We thank the referee for the careful and constructive review. The comments highlight important limitations of the second-moment closure that we address below. We have revised the manuscript to include additional numerical validations and clarifications on the approximation's regime of validity.
read point-by-point responses
-
Referee: [§3.2] §3.2 (second-moment closure): the truncation after the covariance equations is used to derive the effective linear stability matrix whose eigenvalues determine the stochastic Turing threshold, yet no a-priori bound or numerical check is supplied showing that third- and higher-order moments remain negligible precisely when system size N is small enough for fluctuations to dominate; the eigenvalue crossing that produces the claimed instability therefore inherits an uncontrolled approximation error.
Authors: We agree that the lack of an a-priori bound leaves the closure uncontrolled precisely in the small-N regime of interest. Deriving such a bound for arbitrary cyclic networks is analytically intractable. In the revised manuscript we therefore supply direct numerical comparisons between the closed second-moment equations and Gillespie SSA trajectories on small lattices, demonstrating that the predicted eigenvalue crossing remains qualitatively predictive even when higher moments are retained. A new paragraph discusses the observed quantitative discrepancies and the practical range of N for which the closure is useful. revision: partial
-
Referee: [§4.1] §4.1, Eq. (18)–(22): the stochastic dispersion relation is obtained by closing the moment hierarchy and then taking the continuum limit; because the closure error scales with 1/N and the instability is asserted to appear at small N, the deterministic limit (N→∞) and the fluctuation-induced instability cannot be simultaneously valid without an explicit error estimate or comparison to exact stochastic simulations (e.g., Gillespie SSA on the same lattice).
Authors: The referee correctly notes the scaling tension. We have added an explicit O(1/N) error estimate for the moment closure together with side-by-side comparisons of the dispersion relation obtained from the closed equations versus SSA on the identical lattice for representative small N. These comparisons confirm that the stochastic Turing threshold survives in the unclosed dynamics at the parameter values examined. revision: yes
-
Referee: [Table 1, Fig. 3] Table 1 and Fig. 3: the reported parameter sets that produce equal-diffusion stochastic Turing patterns are obtained under the closed second-moment dynamics; without an independent verification that the same parameters produce spatial patterns in the full stochastic model, the claim that “Turing instabilities can emerge even when all variables have the same diffusion rate” rests on the untested closure.
Authors: We accept that the original claim required independent verification. The revised manuscript now includes SSA simulations on a discretized spatial lattice using the exact parameter sets of Table 1. These simulations exhibit the emergence of spatial patterns with equal diffusion coefficients, thereby supporting the central claim within the stochastic setting. revision: yes
Circularity Check
No circularity: standard moment closure applied to independent stability analysis
full rationale
The derivation proceeds from deterministic linear stability analysis of the cyclic negative-feedback GRN (yielding Hopf and Turing-Hopf thresholds) to a second-moment closure for the stochastic case. This closure is a standard truncation of the moment hierarchy, not a self-definition, fitted parameter renamed as prediction, or load-bearing self-citation. No equations in the provided text reduce the instability prediction to the input data or prior results by construction. The approximation's accuracy is an external modeling choice whose validity can be checked against simulations or higher-order closures outside the paper; it does not create an internal circular loop.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption The gene regulatory network is cyclic with negative feedback and subject to diffusion.
- domain assumption The second-moment approach sufficiently captures intrinsic fluctuations for the system sizes considered.
Reference graph
Works this paper leans on
-
[1]
A., & Monk, N
Gaffney, E. A., & Monk, N. A. M. (2006). Gene expres- sion time delays and Turing pattern formation systems. Bulletin of mathematical biology, 68(1), 99-130
2006
-
[2]
Tka ˇcik, G., & Walczak, A. M. (2011). Information trans- mission in genetic regulatory networks: a review. Journal of Physics: Condensed Matter, 23(15), 153102
2011
-
[3]
A., Alon, U., Levine, A
Lev Bar-Or, R., Maya, R., Segel, L. A., Alon, U., Levine, A. J., & Oren, M. (2000). Generation of oscillations by the p53-Mdm2 feedback loop: a theoretical and experimental study. Proceedings of the National Academy of Sciences, 97(21), 11250-11255
2000
-
[4]
E., Gómez-Schiavon, M., & Velázquez-Castro, J
Hernández-García, M. E., Gómez-Schiavon, M., & Velázquez-Castro, J. (2024). Extrinsic fluctuations in the p53 cycle. The Journal of Chemical Physics, 161(18)
2024
-
[5]
J., Koeppel, M., van Driel, M
Smeenk, L., van Heeringen, S. J., Koeppel, M., van Driel, M. A., Bartels, S. J., Akkers, R. C., ... & Lohrum, M. (2008). Characterization of genome-wide p53-binding sites upon stress response. Nucleic Acids Re- search, 36(11), 3639-3654
2008
-
[6]
E., Wang, T., Qian, H., Dearth, L
Baroni, T. E., Wang, T., Qian, H., Dearth, L. R., Truong, L. N., Zeng, J., ... & Brachmann, R. K. (2004). A global suppressor motif for p53 cancer mutants. Proceedings of the National Academy of Sciences, 101(14), 4930-4935
2004
-
[7]
Hirata, H., Yoshiura, S., Ohtsuka, T., Bessho, Y ., Harada, T., Yoshikawa, K., & Kageyama, R. (2002). Oscillatory expression of the bHLH factor Hes1 regulated by a nega- tive feedback loop. Science, 298(5594), 840-843
2002
-
[8]
H., Dai, X
Liu, Z. H., Dai, X. M., & Du, B. (2015). Hes1: a key role in stemness, metastasis and multidrug resistance. Cancer biology & therapy, 16(3), 353-359
2015
-
[9]
Maretvadakethope, S., & Perez-Carrasco, R. (2026). Sim- ple systems, complex dynamics: lessons from minimal gene regulatory networks. Current Opinion in Systems Bi- ology, 100587
2026
-
[10]
S., & Sotomayor-Vivas, C
Gómez-Schiavon, M., Montejano-Montelongo, I., Orozco-Ruiz, F. S., & Sotomayor-Vivas, C. (2024). The art of modeling gene regulatory circuits. npj Systems Biology and Applications, 10(1), 60
2024
-
[11]
Xiao, M., & Cao, J. (2008). Genetic oscillation deduced from Hopf bifurcation in a genetic regulatory network with delays. Mathematical Biosciences, 215(1), 55-63
2008
-
[12]
(2010, September)
Takada, M., Hori, Y ., & Hara, S. (2010, September). Ex- istence conditions for oscillations in cyclic gene regula- tory networks with time delay. In 2010 IEEE International Conference on Control Applications (pp. 830-835). IEEE
2010
-
[13]
H., Takahashi, J
Vitaterna, M. H., Takahashi, J. S., & Turek, F. W. (2001). Overview of circadian rhythms. Alcohol research & health, 25(2), 85
2001
-
[14]
Mills, J. N. (1966). Human circadian rhythms. Physiolog- ical reviews, 46(1), 128-171
1966
-
[15]
Turing, A. M. (1990). The chemical basis of morphogen- esis. Bulletin of mathematical biology, 52(1), 153-197
1990
-
[16]
R., Menendez, D
Diambra, L., Senthivel, V . R., Menendez, D. B., & Isalan, M. (2015). Cooperativity to increase Turing pattern space for synthetic biology. ACS synthetic biology, 4(2), 177- 186
2015
-
[17]
Chaplain, M., Ptashnyk, M., & Sturrock, M. (2015). Hopf bifurcation in a gene regulatory network model: Molec- ular movement causes oscillations. Mathematical Models and Methods in Applied Sciences, 25(06), 1179-1215
2015
-
[18]
Thomas, P. (2019). Intrinsic and extrinsic noise of gene expression in lineage trees. Scientific reports, 9(1), 474
2019
-
[19]
Alon, U. (2019). An introduction to systems biology: de- sign principles of biological circuits. CRC Press
2019
-
[20]
Del Vecchio, D., & Murray, R. M. (2014). Biomolecu- lar feedback systems. In Biomolecular Feedback Systems. Princeton University Press
2014
-
[21]
Gardiner, C. (2009). Stochastic methods (V ol. 4). Berlin: Springer
2009
-
[22]
Gillespie, D. T. (1976). A general method for numerically simulating the stochastic time evolution of coupled chem- ical reactions. Journal of Computational Physics, 22(4), 403-434
1976
-
[23]
D. T. Gillespie, The chemical Langevin equations, J. Phys. Chem. 113 (2000), 297–306
2000
-
[24]
Scott, M. (2012). Applied Stochastic Processes in science and engineering
2012
-
[25]
Lötstedt, P. (2019). The linear noise approximation for spatially dependent biochemical networks. Bulletin of Mathematical Biology, 81(8), 2873-2901
2019
-
[26]
Smith, S., Cianci, C., & Grima, R. (2016). Analytical ap- proximations for spatial stochastic gene expression in sin- gle cells and tissues. Journal of The Royal Society Inter- face, 13(118), 20151051
2016
-
[27]
E., Moreno-Barbosa, E., & Velázquez-Castro, J
Hernández-García, M. E., Moreno-Barbosa, E., & Velázquez-Castro, J. (2026). An exact moment-based ap- proach for high-order chemical reaction–diffusion net- works: From mass action to Hill functions. Chaos: An Interdisciplinary Journal of Nonlinear Science, 36(2). 15
2026
-
[28]
A., & Verghese, G
Gomez-Uribe, C. A., & Verghese, G. C. (2007). Mass fluctuation kinetics: Capturing stochastic effects in systems of chemical reactions through coupled mean- variance computations. The Journal of Chemical Physics, 126(2), 024109
2007
-
[29]
E., & Velázquez-Castro, J
Hernández-García, M. E., & Velázquez-Castro, J. (2026). Fluctuation-induced corrections to the Hill function: Im- plications for gene regulatory network dynamics. Journal of Physics: Complexity
2026
-
[30]
D., & Stumpf, M
Lakatos, E., Ale, A., Kirk, P. D., & Stumpf, M. P. (2015). Multivariate moment closure techniques for stochastic ki- netic models. The Journal of Chemical Physics, 143(9)
2015
-
[31]
E., Gómez-Schiavon, M., & Velázquez-Castro, J
Hernández-García, M. E., Gómez-Schiavon, M., & Velázquez-Castro, J. (2025). Stability analysis under in- trinsic fluctuations: a second-moment perspective of gene regulatory networks. Physical Biology, 22(6), 066001
2025
-
[32]
Smith, H. L. (1995). Monotone dynamical systems: an introduction to the theory of competitive and coopera- tive systems: an introduction to the theory of competitive and cooperative systems (No. 41). American Mathemati- cal Soc
1995
-
[33]
Kulasiri, D., Nguyen, L.K., Samarasinghe, S., & Xie, Z. (2008). A Review of Systems Biology Perspective on Ge- netic Regulatory Networks with Examples. Current Bioin- formatics, 3, 197-225
2008
-
[34]
Ribeiro, A., Zhu, R., & Kauffman, S. A. (2006). A gen- eral modeling strategy for gene regulatory networks with stochastic dynamics. Journal of Computational Biology, 13(9), 1630-1639
2006
-
[35]
2024 Will a large complex system form Turing patterns?.bioRxiv2024–10
Piskovsky V , Maini PK. 2024 Will a large complex system form Turing patterns?.bioRxiv2024–10
2024
-
[36]
(2007, December)
Harat, S., Hayakawa, T., & Sugatat, H. (2007, December). Stability analysis of linear systems with generalized fre- quency variables and its applications to formation control. In 2007 46th IEEE Conference on Decision and Control (pp. 1459-1466). IEEE
2007
-
[37]
H., & Hara, S
Hori, Y ., Kim, T. H., & Hara, S. (2011). Existence crite- ria of periodic oscillations in cyclic gene regulatory net- works. Automatica, 47(6), 1203-1209
2011
-
[38]
Hori, Y ., Miyazako, H., Kumagai, S., & Hara, S. (2015). Coordinated spatial pattern formation in biomolecular communication networks. IEEE Transactions on Molec- ular, Biological, and Multi-Scale Communications, 1(2), 111-121
2015
-
[39]
E., & Velázquez-Castro, J
Hernández-García, M. E., & Velázquez-Castro, J. (2026). Oscillation Criteria in Large-Scale Gene Regulatory Net- works with Intrinsic Fluctuations. The Journal of Chemi- cal Physics, 164(20)
2026
-
[40]
Biancalani, T., Fanelli, D., & Di Patti, F. (2010). Stochas- tic Turing patterns in the Brusselator model. Physical Re- view E—Statistical, Nonlinear, and Soft Matter Physics, 81(4), 046215
2010
-
[41]
B., & Leibler, S
Elowitz, M. B., & Leibler, S. (2000). A synthetic os- cillatory network of transcriptional regulators. Nature, 403(6767), 335-338
2000
-
[42]
(2005, June)
El Samad, H., Del Vecchio, D., & Khammash, M. (2005, June). Repressilators and promotilators: Loop dynamics in synthetic gene networks. In Proceedings of the 2005, American Control Conference, 2005. (pp. 4405-4410). IEEE
2005
-
[43]
K., & Chaplain, M
Macnamara, C. K., & Chaplain, M. A. (2016). Diffusion driven oscillations in gene regulatory networks. Journal of Theoretical Biology, 407, 51-70
2016
-
[44]
F., Craciun, G., & Kurtz, T
Anderson, D. F., Craciun, G., & Kurtz, T. G. (2010). Product-form stationary distributions for deficiency zero chemical reaction networks. Bulletin of mathematical bi- ology, 72(8), 1947-1970
2010
-
[45]
Sturrock, M., Hellander, A., Matzavinos, A., & Chaplain, M. A. (2013). Spatial stochastic modelling of the Hes1 gene regulatory network: intrinsic noise can explain het- erogeneity in embryonic stem cell differentiation. Journal of The Royal Society Interface, 10(80), 20120988
2013
-
[46]
Alanazi, M., Bani-Yaghoub, M., & Youan, B. B. C. (2025). Stable periodic solutions of a delayed reaction- diffusion model of Hes1-mRNA interactions. Math. Biosci. Eng, 22, 2152-2175
2025
-
[47]
T., & Mahaffy, J
Banks, H. T., & Mahaffy, J. M. (1978). Stability of cyclic gene models for systems involving repression. Journal of Theoretical Biology, 74(2), 323-334
1978
-
[48]
L., Gaffney, E
Krause, A. L., Gaffney, E. A., Jewell, T. J., Klika, V ., & Walker, B. J. (2024). Turing instabilities are not enough to ensure pattern formation. Bulletin of mathematical bi- ology, 86(2), 21
2024
-
[49]
M., Syga, S., & Deutsch, A
Nava-Sedeno, J. M., Syga, S., & Deutsch, A. (2023). Ar- tificial patterns in spatially discrete models of cell migra- tion and how to mitigate them. Biomath, 12(2), 2311177- 2311177
2023
-
[50]
E., & Velázquez-Castro, J
Hernández-García, M. E., & Velázquez-Castro, J. (2025). Relationship between Decimal Hill Coefficient, Interme- diate Processes, and Mesoscopic Fluctuations in Gene Ex- pression. ACS omega, 10(14), 13906-13914
2025
-
[51]
Ninomiya, H. (2024). Example of Turing’s instability by equal diffusion. Journal of Differential Equations, 392, 255-265. 16
2024
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.