Co-condensation and multivalency enable acetylation-sensitive, concentration-robust assembly of BRD4 condensates
Pith reviewed 2026-06-28 11:44 UTC · model grok-4.3
The pith
Co-condensation with chromatin lets BRD4 assemble below bulk concentrations while sharpening acetylation sensitivity.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
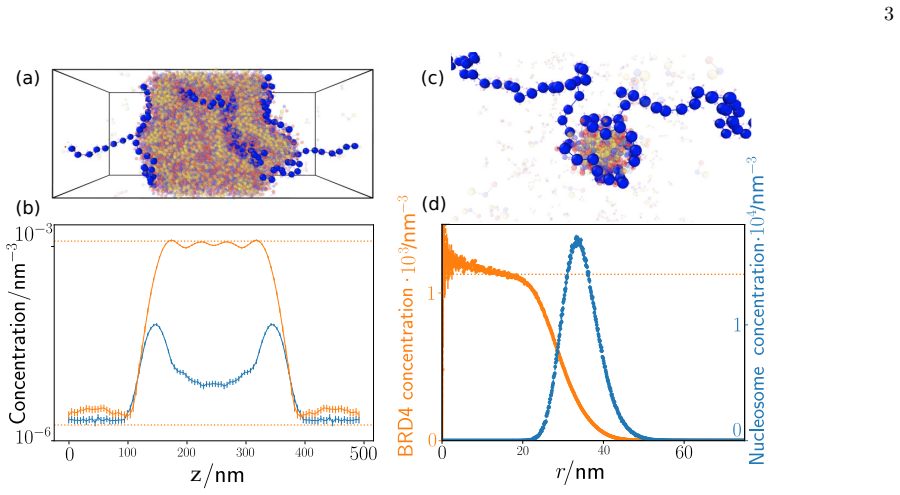
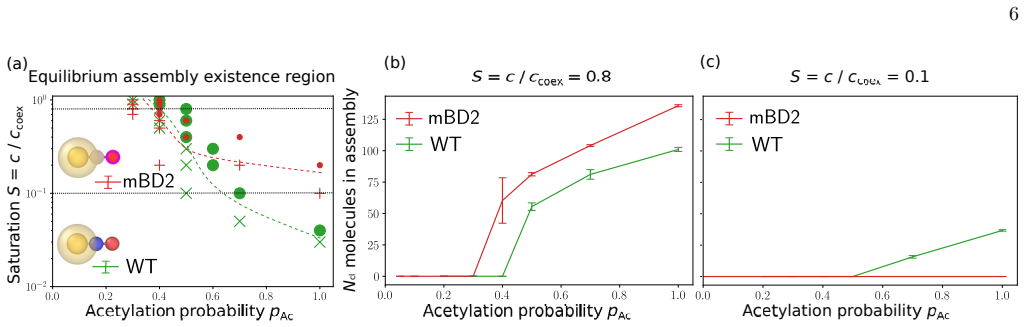
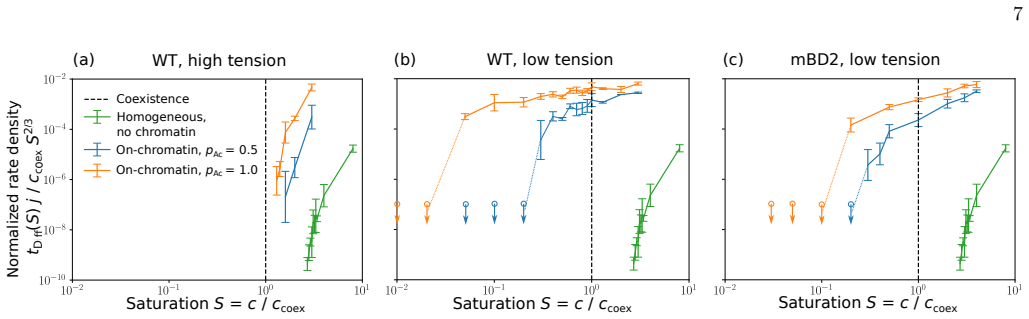
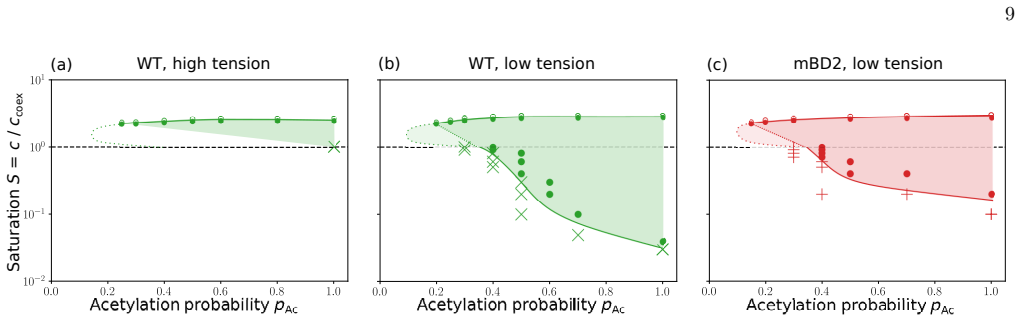
Using an ultra-coarse-grained molecular-dynamics model, we show that co-condensation of BRD4 with chromatin enables rapid assembly below the bulk coexistence concentration, thereby suppressing off-chromatin condensation and enhancing spatial selectivity. Multivalent binding between BRD4 and acetylated histone tails sharpens the dependence of co-condensation on acetylation density through combinatorial effects, increasing contrast between highly acetylated regions and weakly acetylated background chromatin.
What carries the argument
Co-condensation of BRD4 with chromatin enabled by multivalent binding to acetylated histone tails
If this is right
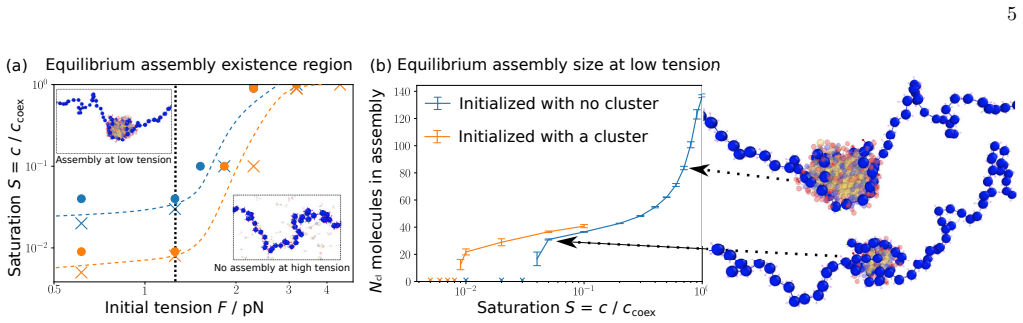
- BRD4 can form condensates at chromatin sites at concentrations too low for phase separation in solution.
- Off-chromatin regions remain free of condensates due to the higher threshold required there.
- The response to acetylation is sharpened, allowing better distinction between modified and unmodified chromatin.
- This provides a way to achieve both sensitivity to chemical marks and robustness to concentration fluctuations.
Where Pith is reading between the lines
- Similar co-condensation mechanisms may operate in other chromatin-binding proteins to achieve precise localization.
- The model could be tested by varying chromatin fiber geometry to see if the robustness holds.
- This might connect to how cells regulate condensate formation during gene expression changes.
Load-bearing premise
The ultra-coarse-grained molecular-dynamics model accurately captures the interaction strengths, binding valences, and chromatin structure relevant to BRD4 condensation.
What would settle it
Direct measurement showing that BRD4 chromatin co-condensation does not occur below the bulk coexistence concentration or that multivalency does not increase acetylation sensitivity.
Figures


read the original abstract
Biomolecular condensates must assemble at specific locations and times inside living cells to perform their biological functions. However, it remains unclear how condensate formation achieves high spatiotemporal precision, responding sensitively to local chemical modifications while remaining robust to fluctuations in protein concentration. Here we study chromatin-associated BRD4 condensates to identify a physical mechanism that enables this combination of sensitivity and robustness. Using an ultra-coarse-grained molecular-dynamics model, we show that co-condensation of BRD4 with chromatin enables rapid assembly below the bulk coexistence concentration, thereby suppressing off-chromatin condensation and enhancing spatial selectivity. Multivalent binding between BRD4 and acetylated histone tails sharpens the dependence of co-condensation on acetylation density through combinatorial effects, increasing contrast between highly acetylated regions and weakly acetylated background chromatin. This mechanism explains how co-condensation and multivalent binding jointly enable sensitive yet robust spatiotemporal targeting by chromatin-associated condensates.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript uses an ultra-coarse-grained molecular-dynamics model of BRD4 to argue that co-condensation with chromatin permits rapid condensate assembly below the bulk coexistence concentration (thereby suppressing off-chromatin droplets and improving spatial selectivity) and that multivalent BRD4–acetylated-histone-tail interactions combinatorially sharpen the response to acetylation density, increasing contrast between modified and background chromatin.
Significance. If the model faithfully reproduces the relevant interaction valences, affinities, and polymer statistics, the work supplies a concrete physical mechanism that reconciles the competing requirements of chemical sensitivity and concentration robustness for chromatin-associated condensates; the combinatorial sharpening effect is a falsifiable prediction that could be tested by varying bromodomain copy number or acetylation density in cells.
major comments (2)
- [Methods] Methods (model parameterization): the interaction strengths and valences between BRD4 and chromatin / acetylated tails are free parameters whose values, calibration procedure, and sensitivity to variation are not reported; because every quantitative claim (sub-bulk assembly threshold, acetylation contrast, concentration robustness) is generated by this model, the absence of validation against measured bromodomain–histone affinities or nucleosome spacing undermines the central conclusions.
- [Results] Results (co-condensation threshold): the reported ability to assemble below bulk coexistence is shown only for the chosen ultra-coarse-grained potentials; without an explicit comparison to a finer-grained or experimental reference system, it remains unclear whether the effect survives changes in effective valence or chromatin persistence length.
minor comments (2)
- [Figures] Figure legends should state the number of independent simulation replicas and the precise definition of the order parameter used to detect condensate formation.
- [Main text] Notation for the acetylation density variable is introduced without an equation; adding a short definition in the main text would improve readability.
Simulated Author's Rebuttal
We thank the referee for the constructive comments, which highlight important aspects of model transparency and robustness. We address each major point below and will revise the manuscript to strengthen the presentation of the ultra-coarse-grained model.
read point-by-point responses
-
Referee: [Methods] Methods (model parameterization): the interaction strengths and valences between BRD4 and chromatin / acetylated tails are free parameters whose values, calibration procedure, and sensitivity to variation are not reported; because every quantitative claim (sub-bulk assembly threshold, acetylation contrast, concentration robustness) is generated by this model, the absence of validation against measured bromodomain–histone affinities or nucleosome spacing undermines the central conclusions.
Authors: We agree that the manuscript should report the specific interaction parameters, their relation to literature values for bromodomain–histone tail affinities, and sensitivity tests. In the revised version we will add a Methods subsection that lists the chosen interaction strengths and valences, cites measured Kd values from the bromodomain literature to justify the order of magnitude, and presents new simulations in which these parameters are varied by ±30 %. These additional runs confirm that the sub-bulk co-condensation threshold and the sharpening of acetylation contrast remain qualitatively intact across the tested range, although the precise numerical thresholds shift. We therefore view the central mechanistic conclusions as robust but acknowledge that the original submission lacked this documentation. revision: yes
-
Referee: [Results] Results (co-condensation threshold): the reported ability to assemble below bulk coexistence is shown only for the chosen ultra-coarse-grained potentials; without an explicit comparison to a finer-grained or experimental reference system, it remains unclear whether the effect survives changes in effective valence or chromatin persistence length.
Authors: The ultra-coarse-grained representation was chosen to access the relevant system sizes and timescales. To address the concern, the revised manuscript will include a new supplementary figure that systematically varies chromatin persistence length (by changing the bending modulus) and effective valence (by altering the number of binding sites per nucleosome). These tests show that the ability to assemble below the bulk coexistence concentration persists for persistence lengths between 10 and 50 nm and for valences differing by up to a factor of two, consistent with the polymer-physics expectation that multivalent co-condensation lowers the effective critical concentration. While a direct one-to-one mapping to a finer-grained model is not feasible for the chromosome-scale systems examined, the additional parameter sweeps provide evidence that the reported effect is not an artifact of the specific potential set. revision: yes
Circularity Check
No circularity detected; results emerge from simulation of stated model
full rationale
The paper reports behaviors observed in an ultra-coarse-grained MD model whose interaction rules and chromatin geometry are defined independently of the target outcomes (co-condensation below bulk concentration, acetylation sensitivity via multivalency). No equations, fitted parameters, or self-citations are invoked such that any reported prediction reduces to a definition or to a quantity chosen to produce the result. The model is presented as an exploratory tool whose fidelity is an assumption, not a tautology; the derivation chain therefore remains self-contained against external benchmarks.
Axiom & Free-Parameter Ledger
free parameters (1)
- BRD4-chromatin and BRD4-acetyl-tail interaction strengths and valences
axioms (1)
- domain assumption The ultra-coarse-grained representation preserves the essential physics of multivalent binding and co-condensation with chromatin.
Reference graph
Works this paper leans on
-
[1]
Y. Xing, C. V. Johnson, P. T. Moen, J. A. McNeil, and J. Lawrence, Nonrandom gene organization: structural arrangements of specific pre-mRNA transcription and splicing with sc-35 domains., J. Cell Biol.131, 1635–1647 10 (1995)
1995
-
[2]
Y. Xing, C. V. Johnson, P. R. Dobner, and J. B. Lawrence, Higher level organization of individual gene transcription and RNA splicing, Science259, 1326–1330 (1993)
1993
-
[3]
P. T. Moen, C. V. Johnson, M. Byron, L. S. Shopland, I. L. de la Serna, A. N. Imbalzano, and J. B. Lawrence, Repositioning of muscle-specific genes relative to the pe- riphery of sc-35 domains during skeletal myogenesis, Mol. Biol. Cell15, 197–206 (2004)
2004
-
[4]
J. M. Brown, J. Green, R. P. das Neves, H. A. Wallace, A. J. Smith, J. Hughes, N. Gray, S. Taylor, W. G. Wood, D. R. Higgs, F. J. Iborra, and V. J. Buckle, Association between active genes occurs at nuclear speckles and is modulated by chromatin environment, J. Cell Biol.182, 1083–1097 (2008)
2008
-
[5]
Boisvert, S
F.-M. Boisvert, S. van Koningsbruggen, J. Navascu´ es, and A. I. Lamond, The multifunctional nucleolus, Nat. Rev. Mol. Cell Biol.8, 574–585 (2007)
2007
-
[6]
Falahati, B
H. Falahati, B. Pelham-Webb, S. Blythe, and E. Wi- eschaus, Nucleation by rRNA dictates the precision of nucleolus assembly, Curr. Biol.26, 277–285 (2016)
2016
-
[7]
Boija, I
A. Boija, I. A. Klein, B. R. Sabari, A. Dall’Agnese, E. L. Coffey, A. V. Zamudio, C. H. Li, K. Shrinivas, J. C. Man- teiga, N. M. Hannett, B. J. Abraham, L. K. Afeyan, Y. E. Guo, J. K. Rimel, C. B. Fant, J. Schuijers, T. I. Lee, D. J. Taatjes, and R. A. Young, Transcription factors activate genes through the phase-separation capacity of their ac- tivation...
2018
-
[8]
B. R. Sabari, A. Dall’Agnese, A. Boija, I. A. Klein, E. L. Coffey, K. Shrinivas, B. J. Abraham, N. M. Hannett, A. V. Zamudio, J. C. Manteiga, C. H. Li, Y. E. Guo, D. S. Day, J. Schuijers, E. Vasile, S. Malik, D. Hnisz, T. I. Lee, I. I. Cisse, R. G. Roeder, P. A. Sharp, A. K. Chakraborty, and R. A. Young, Coactivator condensa- tion at super-enhancers links...
2018
-
[9]
L. Ma, Z. Gao, J. Wu, B. Zhong, Y. Xie, W. Huang, and Y. Lin, Co-condensation between transcription fac- tor and coactivator p300 modulates transcriptional burst- ing kinetics, Mol. Cell81, 1682 (2021)
2021
-
[10]
Zhang, K
Y. Zhang, K. Brown, Y. Yu, Z. Ibrahim, M. Zandian, H. Xuan, S. Ingersoll, T. Lee, C. C. Ebmeier, J. Liu, D. Panne, X. Shi, X. Ren, and T. G. Kutateladze, Nu- clear condensates of p300 formed though the structured catalytic core can act as a storage pool of p300 with re- duced hat activity, Nat. Commun.12(2021)
2021
-
[11]
Lyons, R
H. Lyons, R. T. Veettil, P. Pradhan, C. Fornero, N. De La Cruz, K. Ito, M. Eppert, R. G. Roeder, and B. R. Sabari, Functional partitioning of transcriptional regulators by patterned charge blocks, Cell186, 327 (2023)
2023
-
[12]
Patil, A
A. Patil, A. R. Strom, J. A. Paulo, C. K. Collings, K. M. Ruff, M. K. Shinn, A. Sankar, K. S. Cervantes, T. Wauer, J. D. St. Laurent, G. Xu, L. A. Becker, S. P. Gygi, R. V. Pappu, C. P. Brangwynne, and C. Kadoch, A disordered region controls cbaf activity via condensation and partner recruitment, Cell186, 4936 (2023)
2023
-
[13]
W. F. Richter, S. Nayak, J. Iwasa, and D. J. Taatjes, The mediator complex as a master regulator of transcrip- tion by RNA polymerase ii, Nat. Rev. Mol. Cell Biol.23, 732–749 (2022)
2022
-
[14]
Cho, J.-H
W.-K. Cho, J.-H. Spille, M. Hecht, C. Lee, C. Li, V. Grube, and I. I. Cisse, Mediator and RNA polymerase ii clusters associate in transcription-dependent conden- sates, Science361, 412–415 (2018)
2018
-
[15]
B. L. Crowe, R. C. Larue, C. Yuan, S. Hess, M. Kvaratskhelia, and M. P. Foster, Structure of the brd4 et domain bound to a c-terminal motif fromγ-retroviral integrases reveals a conserved mechanism of interaction, Proc. Natl. Acad. Sci. U.S.A.113, 2086–2091 (2016)
2086
-
[16]
A. R. Strom, J. M. Eeftens, Y. Polyachenko, C. J. Weaver, H.-F. Watanabe, D. Bracha, N. D. Orlovsky, C. C. Jumper, W. M. Jacobs, and C. P. Brangwynne, In- terplay of condensation and chromatin binding underlies brd4 targeting, Mol. Biol. Cell35(2024)
2024
-
[17]
B. N. Devaiah, C. Case-Borden, A. Gegonne, C. H. Hsu, Q. Chen, D. Meerzaman, A. Dey, K. Ozato, and D. S. Singer, Brd4 is a histone acetyltransferase that evicts nu- cleosomes from chromatin, Nat. Struct. Mol. Biol.23, 540–548 (2016)
2016
-
[18]
A. A. Chernov,Modern Crystallography III — Crystal Growth(Springer Berlin Heidelberg, 1984)
1984
-
[19]
S. F. Shimobayashi, P. Ronceray, D. W. Sanders, M. P. Haataja, and C. P. Brangwynne, Nucleation landscape of biomolecular condensates, Nature599, 503 (2021)
2021
-
[20]
Yanagawa and S
M. Yanagawa and S. F. Shimobayashi, Multi-dimensional condensation of intracellular biomolecules, J. Biochem. 175, 179 (2024)
2024
-
[21]
N. A. Erkamp, M. A. Verwiel, D. Qian, T. Sneideris, F. A. Spaepen, D. A. Weitz, J. C. van Hest, and T. P. Knowles, Biomolecular condensates with complex architectures via controlled nucleation, Nat. Chem. Eng.1, 430 (2024)
2024
-
[22]
Shrinivas, B
K. Shrinivas, B. R. Sabari, E. L. Coffey, I. A. Klein, A. Boija, A. V. Zamudio, J. Schuijers, N. M. Hannett, P. A. Sharp, R. A. Young, and A. K. Chakraborty, En- hancer features that drive formation of transcriptional condensates, Mol. Cell75, 549 (2019)
2019
-
[23]
Sharma, K.-J
R. Sharma, K.-J. Choi, M. D. Quan, S. Sharma, B. Sankaran, H. Park, A. LaGrone, J. J. Kim, K. R. MacKenzie, A. C. M. Ferreon, C. Kim, and J. C. Ferreon, Liquid condensation of reprogramming factor klf4 with DNA provides a mechanism for chromatin organization, Nat. Commun.12(2021)
2021
-
[24]
D. A. Porter and K. E. Easterling,Phase transformations in metals and alloys (revised reprint)(CRC press, 2009)
2009
-
[25]
Quail, S
T. Quail, S. Golfier, M. Elsner, K. Ishihara, V. Muruge- san, R. Renger, F. J¨ ulicher, and J. Brugu´ es, Force gener- ation by protein–DNA co-condensation, Nat. Phys.17, 1007–1012 (2021)
2021
-
[26]
Renger, J
R. Renger, J. A. Morin, R. Lemaitre, M. Ruer- Gruss, F. J¨ ulicher, A. Hermann, and S. W. Grill, Co-condensation of proteins with single- and double- stranded DNA, Proc. Natl. Acad. Sci. U.S.A.119(2022)
2022
-
[27]
M. N. Rouches and B. B. Machta, Protein-DNA co- condensation is prewetting to a collapsed polymer, Bio- phys. J.124, 2280–2290 (2025)
2025
-
[28]
T. Curk, J. Dobnikar, and D. Frenkel, Design princi- ples for super selectivity using multivalent interactions (2017)
2017
-
[29]
Z. Xie, S. Angioletti-Uberti, J. Dobnikar, D. Frenkel, and T. Curk, Receptor clustering tunes and sharpens the se- lectivity of multivalent binding, Proc. Natl. Acad. Sci. U.S.A.122(2025)
2025
-
[30]
X. Xia, Y. Peng, and R. Ni, Entropy-driven physical am- plification in multivalent biosensing (2025)
2025
-
[31]
F. E. Regnier and J. Kim, Proteins and proteoforms: New separation challenges, Anal. Chem.90, 361–373 11 (2017)
2017
-
[32]
Panigrahi and B
A. Panigrahi and B. W. O’Malley, Mechanisms of en- hancer action: the known and the unknown, Genome Biol.22(2021)
2021
-
[33]
K. Wu, D. Fan, H. Zhao, Z. Liu, Z. Hou, W. Tao, G. Yu, S. Yuan, X. Zhu, M. Kang, Y. Tian, Z.-J. Chen, J. Liu, and L. Gao, Dynamics of histone acetylation during hu- man early embryogenesis, Cell Discov.9(2023)
2023
-
[34]
M. D. Litt, Transitions in histone acetylation reveal boundaries of three separately regulated neighboring loci, EMBO J.20, 2224–2235 (2001)
2001
-
[35]
Hensley, W
A. Hensley, W. M. Jacobs, and W. B. Rogers, Self- assembly of photonic crystals by controlling the nucle- ation and growth of DNA-coated colloids, Proc. Natl. Acad. Sci. U.S.A.119, e2114050118 (2022)
2022
-
[36]
Nordenski¨ old, X
L. Nordenski¨ old, X. Shi, N. Korolev, L. Zhao, Z. Zhai, and B. Lindman, Liquid-liquid phase separation (llps) in DNA and chromatin systems from the perspective of col- loid physical chemistry, Adv. Coll. Int. Sci.326, 103133 (2024)
2024
-
[37]
C. A. Brackley, S. Taylor, A. Papantonis, P. R. Cook, and D. Marenduzzo, Nonspecific bridging-induced attraction drives clustering of DNA-binding proteins and genome organization, Proc. Natl. Acad. Sci. U.S.A.110(2013)
2013
-
[38]
J.-K. Ryu, C. Bouchoux, H. W. Liu, E. Kim, M. Mi- namino, R. de Groot, A. J. Katan, A. Bonato, D. Maren- duzzo, D. Michieletto, F. Uhlmann, and C. Dekker, Bridging-induced phase separation induced by cohesin smc protein complexes, Sci. Adv.7(2021)
2021
-
[39]
Chong, C
S. Chong, C. Dugast-Darzacq, Z. Liu, P. Dong, G. M. Dailey, C. Cattoglio, A. Heckert, S. Banala, L. Lavis, X. Darzacq, and R. Tjian, Imaging dynamic and se- lective low-complexity domain interactions that control gene transcription, Science361(2018)
2018
-
[40]
Woringer and X
M. Woringer and X. Darzacq, Protein motion in the nu- cleus: from anomalous diffusion to weak interactions, Biochem. Soc. Trans.46, 945–956 (2018)
2018
-
[41]
Alberti, A
S. Alberti, A. Gladfelter, and T. Mittag, Considerations and challenges in studying liquid-liquid phase separation and biomolecular condensates, Cell176, 419–434 (2019)
2019
-
[42]
G. J. Narlikar, S. Myong, D. Larson, K. Maeshima, N. Francis, K. Rippe, B. Sabari, L. Strader, and R. Tjian, Is transcriptional regulation just going through a phase?, Mol. Cell81, 1579–1585 (2021)
2021
-
[43]
B. R. Sabari, A. A. Hyman, and D. Hnisz, Functional specificity in biomolecular condensates revealed by ge- netic complementation, Nat. Rev. Genet.26, 279–290 (2024)
2024
-
[44]
Bremer, W
A. Bremer, W. H. Lang, R. P. Kempen, K. Sweta, A. B. Taylor, M. B. Borgia, A. Z. Ansari, and T. Mittag, Rec- onciling competing models on the roles of condensates and soluble complexes in transcription factor function, Mol. Cell85, 2718 (2025)
2025
-
[45]
V. B. Fan, A. A. Abidi, T. G. Graham, X. Darzacq, and M. V. Staller, Short activation domains control chromatin association of transcription factors, eLife15 (2026)
2026
-
[46]
D. L. Ermak and H. Buckholz, Numerical integration of the langevin equation: Monte carlo simulation, Journal of Computational Physics35, 169–182 (1980)
1980
-
[47]
Kremer and G
K. Kremer and G. S. Grest, Dynamics of entangled linear polymer melts: A molecular-dynamics simulation, The Journal of Chemical Physics92, 5057–5086 (1990)
1990
-
[48]
Beltran, D
B. Beltran, D. Kannan, Q. MacPherson, and A. J. Spakowitz, Geometrical heterogeneity dominates ther- mal fluctuations in facilitating chromatin contacts, Phys. Rev. Lett.123(2019)
2019
-
[49]
M. A. Karymov, M. Tomschik, S. H. Leuba, P. Caiafa, and J. Zlatanova, DNA methylation-dependent chro- matin fiber compaction in vivo and in vitro: requirement for linker histone, FASEB J.15, 2631–2641 (2001)
2001
-
[50]
Cheng, P
P. Cheng, P. M. Oliver, M. J. Barrett, and D. Vezenov, Progress toward the application of molecular force spec- troscopy to ¡scp¿DNA¡/scp¿ sequencing, Electrophoresis 33, 3497–3505 (2012)
2012
-
[51]
T. J. Richmond, J. T. Finch, B. Rushton, D. Rhodes, and A. Klug, Structure of the nucleosome core particle at 7 ˚A resolution, Nature311, 532–537 (1984)
1984
-
[52]
Jumper, R
J. Jumper, R. Evans, A. Pritzel, T. Green, M. Fig- urnov, O. Ronneberger, K. Tunyasuvunakool, R. Bates, A. ˇZ´ ıdek, A. Potapenko, A. Bridgland, C. Meyer, S. A. A. Kohl, A. J. Ballard, A. Cowie, B. Romera-Paredes, S. Nikolov, R. Jain, J. Adler, T. Back, S. Petersen, D. Reiman, E. Clancy, M. Zielinski, M. Steinegger, M. Pacholska, T. Berghammer, S. Bodenst...
2021
-
[53]
A. A. Zamyatnin, Amino acid, peptide, and protein volume in solution, Annu. Rev. Biophys. Bioeng.13, 145–165 (1984)
1984
-
[54]
Sarkar, S
A. Sarkar, S. Caamano, and J. M. Fernandez, The elas- ticity of individual titin pevk exons measured by single molecule atomic force microscopy, J. Biol. Chem.280, 6261–6264 (2005)
2005
-
[55]
H. P. Erickson, Reversible unfolding of fibronectin type iii and immunoglobulin domains provides the structural basis for stretch and elasticity of titin and fibronectin., Proc. Natl. Acad. Sci. U.S.A.91, 10114–10118 (1994)
1994
-
[56]
Trombit´ as, M
K. Trombit´ as, M. Greaser, S. Labeit, J.-P. Jin, M. Keller- mayer, M. Helmes, and H. Granzier, Titin extensibility in situ: Entropic elasticity of permanently folded and per- manently unfolded molecular segments, J. Cell Biol.140, 853–859 (1998)
1998
-
[57]
P. J. Flory, Statistical mechanics of dilute polymer solu- tions, J. Chem. Phys.17, 1347–1348 (1949)
1949
-
[58]
P. J. Flory and W. R. Krigbaum, Statistical mechan- ics of dilute polymer solutions. ii, J. Chem. Phys.18, 1086–1094 (1950)
1950
-
[59]
soft colloids
A. A. Louis, P. G. Bolhuis, J. P. Hansen, and E. J. Meijer, Can polymer coils be modeled as “soft colloids”?, Phys. Rev. Lett.85, 2522–2525 (2000)
2000
-
[60]
Bracha, M
D. Bracha, M. T. Walls, M.-T. Wei, L. Zhu, M. Kurian, J. L. Avalos, J. E. Toettcher, and C. P. Brangwynne, Mapping local and global liquid phase behavior in liv- ing cells using photo-oligomerizable seeds, Cell175, 1467 (2018)
2018
-
[61]
Vollmuth, W
F. Vollmuth, W. Blankenfeldt, and M. Geyer, Struc- tures of the dual bromodomains of the p-tefb-activating protein brd4 at atomic resolution, J. Biol. Chem.284, 36547–36556 (2009)
2009
-
[62]
Co-condensation and multivalency enable acetylation-sensitive, concentration-robust assembly of BRD4 condensates
A. Dey, F. Chitsaz, A. Abbasi, T. Misteli, and K. Ozato, The double bromodomain protein brd4 binds to acety- lated chromatin during interphase and mitosis, Proc. Natl. Acad. Sci. U.S.A.100, 8758–8763 (2003). 12 Supplementary Information for “Co-condensation and multivalency enable acetylation-sensitive, concentration-robust assembly of BRD4 condensates” S...
2003
-
[63]
Disable steric WCA repulsion between BD domains and acetylated tails, allowing an Ac tail to occupy the center of the binding pocket
-
[64]
The range is chosen as Rbind ≈0.34σ BDAc/2, whereσ BDAc = (σBD +σ Ac)/2, andσ BD andσ Ac are the WCA diameters of a BD bead and an acetylated-tail bead, respectively
Introduce a Gaussian attraction with well depthU bind and a short interaction range. The range is chosen as Rbind ≈0.34σ BDAc/2, whereσ BDAc = (σBD +σ Ac)/2, andσ BD andσ Ac are the WCA diameters of a BD bead and an acetylated-tail bead, respectively. This choice ensures that the attraction is significantly shorter-ranged than the steric exclusion between...
-
[65]
This prevents multiple Ac tails from simultaneously approaching a bound BD domain
Introduce steric repulsion between acetylated tails with effective sizeσAcAc ≈0.85σ BDBD. This prevents multiple Ac tails from simultaneously approaching a bound BD domain. The value is chosen as a compromise, as it is large enough to enforce single occupancy, but small enough to avoid artificially disrupting droplet structure or histone tail packing
-
[66]
Short linker lengths combined with nonlinear bond potentials would otherwise generate large forces over short distances, requiring prohibitively small integration timesteps
Extend the maximum bond length of histone tails fromℓ IDR/2 to 3ℓIDR/4, whereℓ IDR is the length of the IDR portion of the corresponding histone tail. Short linker lengths combined with nonlinear bond potentials would otherwise generate large forces over short distances, requiring prohibitively small integration timesteps. This extension reduces stiffness...
-
[67]
In the coarse-grained representation, tails would oth- erwise have very small mass, leading to large accelerations under strong, short-range binding forces
Increase the mass of acetylated tails tom Ac ≈0.6m BRD4. In the coarse-grained representation, tails would oth- erwise have very small mass, leading to large accelerations under strong, short-range binding forces. Increasing the mass stabilizes integration while primarily affecting dynamics rather than equilibrium properties, making it the least disruptiv...
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.