Single-morphogen Turing instability driven by nonlinear intracellular-extracellular coupling
Pith reviewed 2026-06-26 14:31 UTC · model grok-4.3
The pith
Compartmentalizing a single molecular species into intracellular and extracellular fields with nonlinear coupling produces Turing instabilities.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
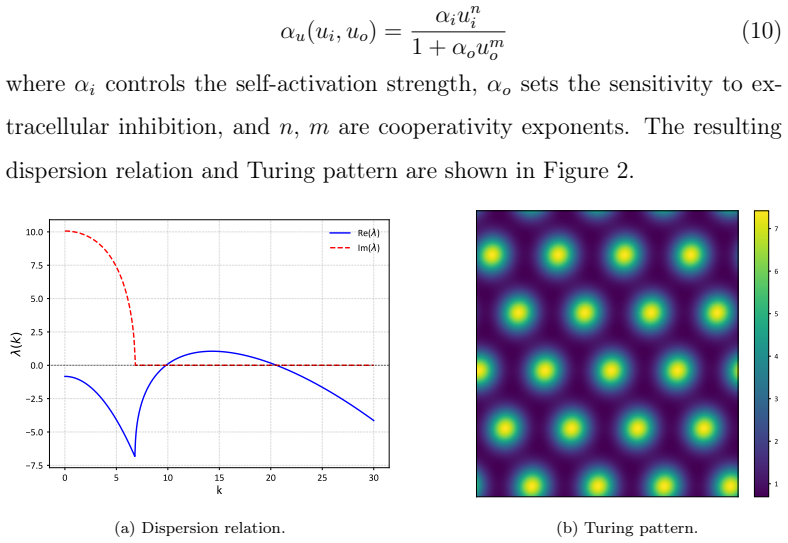
Compartmentalizing a single molecular species into intracellular and extracellular fields, and coupling them through membrane transport or nonlinear basal production rates, can produce diffusion-driven (Turing) instabilities, derived by linearizing the two-field system and verified in three examples with numerical simulations.
What carries the argument
The linearized two-field system of intracellular and extracellular concentrations, whose stability analysis yields the Turing conditions for spatial pattern formation.
If this is right
- Tissue compartmentalization alone can enable pattern formation traditionally attributed to multi-species systems.
- Single-morphogen systems can satisfy the mathematical conditions for diffusion-driven instability.
- Numerical simulations of the three examples confirm that spatial patterns arise when the derived conditions hold.
Where Pith is reading between the lines
- Developmental models might be reduced by replacing multi-species networks with compartmental single-species dynamics.
- Synthetic circuits could be engineered to test pattern formation using only one species plus membrane nonlinearity.
- The same compartmental mechanism may apply to non-biological reaction-diffusion systems that have natural spatial divisions.
Load-bearing premise
The nonlinear coupling terms are such that the linearized two-field system satisfies the derived Turing instability conditions.
What would settle it
A system with compartmentalized single-species concentrations and the specified nonlinear couplings where linear stability analysis predicts no instability yet spatial patterns still appear, or where analysis predicts instability but no patterns form in simulation or experiment.
Figures


read the original abstract
We show that compartmentalizing a single molecular species into intracellular and extracellular fields, and coupling them through membrane transport or nonlinear basal production rates, can produce diffusion-driven (Turing) instabilities. By linearizing the two-field system, we derive the corresponding Turing conditions under which such instabilities may arise. We present three biologically motivated examples that satisfy these conditions and demonstrate the resulting spatial patterns through numerical simulations. These results indicate that tissue compartmentalization alone might enable pattern formation traditionally attributed to multi-species systems.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript claims that compartmentalizing a single molecular species into intracellular (u) and extracellular (v) fields, coupled nonlinearly via membrane transport or basal production rates, can produce diffusion-driven Turing instabilities in a two-field reaction-diffusion system. The authors derive the corresponding instability conditions by linearizing the system around a homogeneous steady state, present three biologically motivated nonlinear coupling examples that satisfy the conditions (negative trace and positive determinant of the reaction Jacobian without diffusion, but wavenumber-dependent instability with diffusion), and demonstrate the resulting spatial patterns via numerical simulations. This indicates that tissue compartmentalization alone might enable pattern formation traditionally requiring multiple species.
Significance. If the central claim holds, the result is significant for developmental biology because it shows how a single morphogen can generate Turing patterns through compartmentalization and nonlinear coupling, potentially reducing the number of required species in models of tissue patterning. The authors receive credit for deriving explicit Turing conditions on the Jacobian, supplying three concrete examples that satisfy them, and providing supporting numerical simulations that produce the expected patterns; these elements make the claim directly testable rather than purely theoretical.
major comments (2)
- [§4] §4 (examples): the manuscript states that the three nonlinear coupling forms satisfy the derived Turing conditions, but does not display the explicit Jacobian matrices or the algebraic verification that trace(J) < 0 and det(J) > 0 hold while the diffusion term allows instability; this verification is load-bearing for the claim that the examples work.
- [Numerical results section] Numerical results section: the simulations are presented as confirming the instability, yet no comparison is given between the analytically predicted growth rate (from the dispersion relation) and the early-time exponential growth observed in the simulations for the chosen parameters; without this check the nonlinear regime could deviate from the linear prediction.
minor comments (3)
- [Abstract] Abstract: the phrase 'single-morphogen' is used while the system is two-field; a parenthetical clarification that the morphogen is compartmentalized would avoid potential misreading.
- Equation numbering: several reaction terms and the dispersion relation are presented without numbers, making cross-reference in the text and in the examples difficult.
- Figure captions: the captions for the simulation figures do not list the exact parameter values used, which would aid reproducibility.
Simulated Author's Rebuttal
We thank the referee for their careful reading of the manuscript and for the constructive comments. We address each major point below and have revised the manuscript accordingly to improve clarity and strengthen the presentation of the results.
read point-by-point responses
-
Referee: §4 (examples): the manuscript states that the three nonlinear coupling forms satisfy the derived Turing conditions, but does not display the explicit Jacobian matrices or the algebraic verification that trace(J) < 0 and det(J) > 0 hold while the diffusion term allows instability; this verification is load-bearing for the claim that the examples work.
Authors: We agree that including the explicit Jacobian matrices and the step-by-step algebraic verification would enhance transparency and make the satisfaction of the Turing conditions more readily verifiable. In the revised version we have added the full Jacobian matrix for each of the three biologically motivated coupling examples, together with the explicit calculations confirming trace(J) < 0, det(J) > 0 in the absence of diffusion and the wavenumber-dependent instability condition once diffusion is included. revision: yes
-
Referee: Numerical results section: the simulations are presented as confirming the instability, yet no comparison is given between the analytically predicted growth rate (from the dispersion relation) and the early-time exponential growth observed in the simulations for the chosen parameters; without this check the nonlinear regime could deviate from the linear prediction.
Authors: We accept that a direct quantitative comparison between the linear dispersion relation and the early-time simulation growth would provide stronger validation. We have therefore added a supplementary panel (or subsection) that extracts the initial exponential growth rate from the simulations for the chosen parameter sets and overlays it against the analytically predicted maximum growth rate from the dispersion relation, confirming consistency in the linear regime. revision: yes
Circularity Check
No significant circularity; derivation is self-contained
full rationale
The paper performs standard linear stability analysis on a two-field (intracellular u, extracellular v) reaction-diffusion system, derives the general Turing conditions on the Jacobian (negative trace, positive determinant without diffusion, but sign change with diffusion), and then explicitly constructs three nonlinear coupling examples (membrane transport and basal production) that satisfy those inequalities by direct substitution. Numerical simulations confirm pattern formation for those examples. No load-bearing step reduces to a fitted parameter renamed as prediction, no self-citation chain justifies the central premise, and the uniqueness of the result is not imported from prior author work. The derivation chain is therefore independent of its own outputs.
Axiom & Free-Parameter Ledger
Reference graph
Works this paper leans on
-
[1]
Series B, Biological Sciences 237 (641) (1952) 37–72, publisher: Royal Society.doi:10.1098/rstb
A.M.Turing, Thechemicalbasisofmorphogenesis, PhilosophicalTrans- actions of the Royal Society of London. Series B, Biological Sciences 237 (641) (1952) 37–72, publisher: Royal Society.doi:10.1098/rstb. 1952.0012
-
[2]
A. Gierer, H. Meinhardt, A theory of biological pattern formation, Ky- bernetik 12 (1) (1972) 30–39.doi:10.1007/BF00289234
-
[3]
J. D. Murray, Mathematical Biology, Springer, Berlin, Heidelberg, 1989. doi:10.1007/978-3-662-08539-4. 14
-
[4]
S. Kondo, T. Miura, Reaction-Diffusion Model as a Framework for Un- derstanding Biological Pattern Formation, Science 329 (5999) (2010) 1616–1620, publisher: American Association for the Advancement of Science (AAAS).doi:10.1126/science.1179047
-
[5]
S. Kondo, M. Watanabe, S. Miyazawa, Studies of Turing pattern forma- tion in zebrafish skin, Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences 379 (2213) (2021) 20200274, publisher: Royal Society.doi:10.1098/rsta.2020.0274
-
[6]
A petavoxel fragment of human cerebral cortex reconstructed at nanoscale resolution
J. Raspopovic, L. Marcon, L. Russo, J. Sharpe, Digit patterning is con- trolled by a Bmp-Sox9-Wnt Turing network modulated by morphogen gradients, Science 345 (6196) (2014) 566–570.doi:10.1126/science. 1252960
-
[7]
J. R. Mooney, B. N. Nagorcka, Spatial patterns produced by a Reaction- diffusion system in primary hair follicles, Journal of Theoretical Biology 115 (2) (1985) 299–317.doi:10.1016/S0022-5193(85)80102-8
-
[8]
S. Sick, S. Reinker, J. Timmer, T. Schlake, WNT and DKK Determine Hair Follicle Spacing Through a Reaction-Diffusion Mechanism, Science 314 (5804) (2006) 1447–1450.doi:10.1126/science.1130088. URLhttps://www.science.org/doi/abs/10.1126/science. 1130088
-
[9]
A. N. Landge, B. M. Jordan, X. Diego, P. Müller, Pattern formation mechanisms of self-organizing reaction-diffusion systems, Developmental Biology 460 (1) (2020) 2–11.doi:10.1016/j.ydbio.2019.10.031. 15
-
[10]
S. Paul, J. Adetunji, T. Hong, Widespread biochemical reaction net- works enable Turing patterns without imposed feedback, Nature Com- munications 15 (1) (2024) 8380.doi:10.1038/s41467-024-52591-0
-
[11]
L. Szili, J. Tóth, On the origin of Turing instability, Journal of Mathematical Chemistry 22 (1) (1997) 39–53.doi:10.1023/A: 1019159427561
work page doi:10.1023/a: 1997
-
[12]
V. Méndez, S. Fedotov, W. Horsthemke, Reaction–Transport Systems, Springer Series in Synergetics, Springer, Berlin, Heidelberg, 2010.doi: 10.1007/978-3-642-11443-4
-
[13]
B. I. Henry, S. L. Wearne, Fractional reaction-diffusion, Physica a- Statistical Mechanics and Its Applications 276 (3-4) (2000) 448–455, publisher: Elsevier Science.doi:10.1016/S0378-4371(99)00469-0
-
[14]
R. A. Satnoianu, M. Menzinger, P. K. Maini, Turing instabilities in general systems, Journal of Mathematical Biology 41 (6) (2000) 493– 512.doi:10.1007/s002850000056
-
[15]
S. Wang, J. Garcia-Ojalvo, M. B. Elowitz, Periodic spatial patterning with a single morphogen, Cell Systems 13 (12) (2022) 1033–1047.e7. doi:10.1016/j.cels.2022.11.001
-
[16]
P.Recho, A.Hallou, E.Hannezo, Theoryofmechanochemicalpatterning in biphasic biological tissues, Proceedings of the National Academy of Sciences of the United States of America 116 (12) (2019) 5344–5349. doi:10.1073/pnas.1813255116. 16
-
[17]
N. Mahashri, A. L. Krause, M. Chandru, T. E. Woolley, Patterns in Time and Space from a Single Morphogen via Nonlinear Layering, arXiv.org (May 2026). URLhttps://arxiv.org/abs/2605.05063v1
Pith/arXiv arXiv 2026
-
[18]
J. Keener, J. Sneyd, Mathematical Physiology, Vol. 8/1 of Interdisci- plinary Applied Mathematics, Springer, New York, NY, 2009.doi: 10.1007/978-0-387-75847-3
-
[19]
Mean-field models for morphogenetic processes in physiological contexts
D.Hernández, AlejandroValdésLópez, E.C.Herrera-Hernández, Mean- field models for morphogenetic processes in physiological contexts, arXiv:2606.10355 [nlin.PS] (Jun. 2026).doi:10.48550/arXiv.2606. 10355. URLhttp://arxiv.org/abs/2606.10355
work page internal anchor Pith review Pith/arXiv arXiv doi:10.48550/arxiv.2606 2026
-
[20]
T. E. Woolley, A. L. Krause, E. A. Gaffney, Bespoke Turing Systems, Bulletin of Mathematical Biology 83 (5) (2021) 41.doi:10.1007/ s11538-021-00870-y
2021
-
[21]
J. Gudyka, J. Ceja-Vega, K. Ivanchenko, Z. Morocho, M. Panella, A. Gamez Hernandez, C. Clarke, E. Perez, S. Silverberg, S. Lee, Concentration-Dependent Effects of Curcumin on Membrane Perme- ability and Structure, ACS Pharmacology & Translational Science 7 (5) (2024) 1546–1556.doi:10.1021/acsptsci.4c00093
-
[22]
M. Queralt-Martín, L. M. Alvero-González, D. A. Perini, E. García- Giménez, A. Alcaraz, Ion transport in biological ion channels beyond classical electrostatics. Nanoscale confinement, non-linear concentration 17 patterns and interfacial effects, Biophysical Reviews (Feb. 2026).doi: 10.1007/s12551-026-01413-2
-
[23]
W. K. Kim, S. Milster, R. Roa, M. Kanduč, J. Dzubiella, Permeability of Polymer Membranes beyond Linear Response, Macromolecules 55 (16) (2022) 7327–7339.doi:10.1021/acs.macromol.2c00605
-
[24]
S. Milster, W. K. Kim, J. Dzubiella, Feedback-controlled solute trans- port through chemo-responsive polymer membranes, The Journal of Chemical Physics 158 (10) (2023) 104903.doi:10.1063/5.0135707
-
[25]
D. Zwicker, py-pde: A python package for solving partial differential equations, Journal of Open Source Software 5 (48) (2020) 2158.doi: 10.21105/joss.02158
-
[26]
P. K. Maini, T. E. Woolley, R. E. Baker, E. A. Gaffney, S. S. Lee, Tur- ing’s model for biological pattern formation and the robustness prob- lem, Interface Focus 2 (4) (2012) 487–496, publisher: Royal Society. doi:10.1098/rsfs.2011.0113
-
[27]
S. T. Vittadello, T. Leyshon, D. Schnoerr, M. P. H. Stumpf, Turing pattern design principles and their robustness, Philosophical Transac- tions of the Royal Society A: Mathematical, Physical and Engineering Sciences 379 (2213) (2021) 20200272.doi:10.1098/rsta.2020.0272. URLhttps://royalsocietypublishing.org/doi/full/10.1098/ rsta.2020.0272
-
[28]
Y. Enomoto, H. Katsura, T. Fujimura, A. Ogata, S. Baba, A. Yamaoka, M. Kihara, T. Abe, O. Nishimura, M. Kadota, D. Hazama, Y. Tanaka, 18 Y. Maniwa, T. Nagano, M. Morimoto, Autocrine tgf-β-positive feedback in profibrotic at2-lineage cells plays a crucial role in non-inflammatory lung fibrogenesis, Nature Communications 14 (1) (2023) 4956.doi: 10.1038/s414...
-
[29]
C. M. Waters, B. L. Bassler, QUORUM SENSING: Cell-to-Cell Commu- nication in Bacteria, Annual Review of Cell and Developmental Biology 21 (Volume 21, 2005) (2005) 319–346.doi:10.1146/annurev.cellbio. 21.012704.131001
-
[30]
S. Moreno-Gámez, M. E. Hochberg, G. S. van Doorn, Quorum sensing as a mechanism to harness the wisdom of the crowds, Nature Commu- nications 14 (1) (2023) 3415.doi:10.1038/s41467-023-37950-7
-
[31]
B. N. Kholodenko, Cell-signalling dynamics in time and space, Nature Reviews Molecular Cell Biology 7 (3) (2006) 165–176.doi:10.1038/ nrm1838
2006
-
[32]
J. E. Purvis, G. Lahav, Encoding and Decoding Cellular Information through Signaling Dynamics, Cell 152 (5) (2013) 945–956.doi:10. 1016/j.cell.2013.02.005
2013
-
[33]
S. Y. Shvartsman, M. P. Hagan, A. Yacoub, P. Dent, H. S. Wiley, D. A. Lauffenburger, Autocrine loops with positive feedback enable context- dependent cell signaling, American Journal of Physiology-Cell Physiol- ogy 282 (3) (2002) C545–C559, publisher: American Physiological Soci- ety.doi:10.1152/ajpcell.00260.2001. 19
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.