Independent channels for miRNA biosynthesis ensure efficient static and dynamic control in the regulation of the early stages of myogenesis
Pith reviewed 2026-05-25 17:00 UTC · model grok-4.3
The pith
Two independent miRNA biosynthesis channels are both required for proper static and dynamic control during early muscle cell differentiation.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
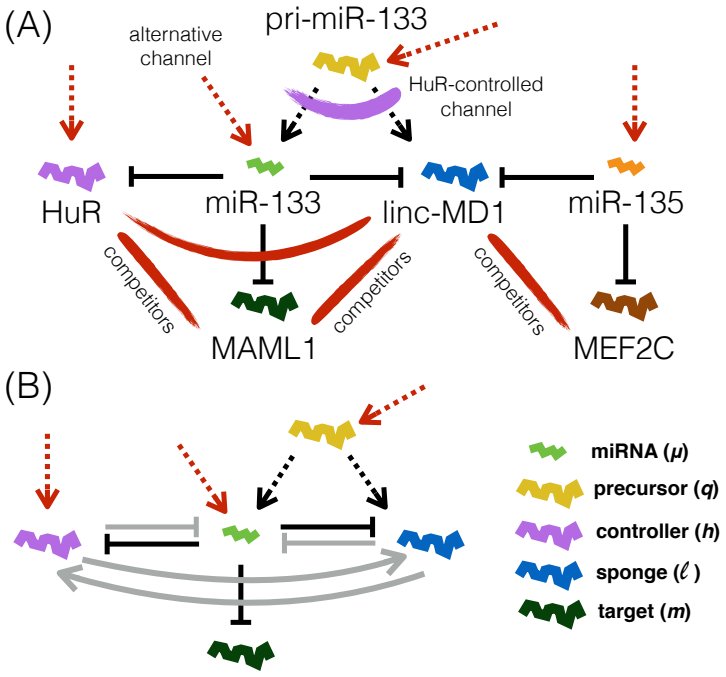
While the miRNA-decoy system can deliver optimal static control, it forces the kinetic parameters into narrow ranges that cross-link the channels; the independent transcription locus supplies the nonlinear target response that permits the fast miRNA concentration changes demanded by differentiation. Static competition-mediated regulation can be achieved by the decoy channel alone, yet both channels prove essential for the circuit's full functionality.
What carries the argument
The dual-channel miRNA architecture in which one channel is a decoy system under exogenous control and the other is direct transcription from a distinct locus, with competition for miRNA binding as the shared control step.
If this is right
- Optimal steady-state control via the decoy system restricts parameters so that the two channels become tightly interdependent.
- Fast concentration shifts during differentiation arise specifically from the nonlinear response of the target to modest increases in the second-channel transcription rate.
- Static regulation can be handled by the decoy channel in isolation, but full circuit performance requires both channels.
- This joint-control pattern may constitute a minimal optimal architecture for miRNA circuits in other regulatory contexts.
Where Pith is reading between the lines
- The same two-channel pattern could be searched for in other differentiation programs that require both precise steady states and timed transitions.
- Knocking out one channel in a cellular model and measuring the separate loss of either precision or timing would directly test the necessity claim.
- The cross-linking of parameters may reduce the need for external tuning by letting one channel compensate for fluctuations in the other.
Load-bearing premise
The two miRNA production channels operate independently and competition for binding is the main control mechanism, so that parameters can be restricted to narrow ranges without needing separate experimental confirmation of those ranges or the independence assumption.
What would settle it
An experiment that removes or blocks the independent genomic transcription locus and still observes both stable target levels and the required rapid miRNA increase during differentiation would falsify the claim that both channels are essential.
Figures






read the original abstract
Motivated by recent experimental work, we define and study a deterministic model of the complex miRNA-based regulatory circuit that putatively controls the early stage of myogenesis in human. We aim in particular at a quantitative understanding of (i) the roles played by the separate and independent miRNA biosynthesis channels (one involving a miRNA-decoy system regulated by an exogenous controller, the other given by transcription from a distinct genomic locus) that appear to be crucial for the differentiation program, and of (ii) how competition to bind miRNAs can efficiently control molecular levels in such an interconnected architecture. We show that optimal static control via the miRNA-decoy system constrains kinetic parameters in narrow ranges where the channels are tightly cross-linked. On the other hand, the alternative locus for miRNA transcription can ensure that the fast concentration shifts required by the differentiation program are achieved, specifically via non-linear response of the target to even modest surges in the miRNA transcription rate. While static, competition-mediated regulation can be achieved by the miRNA-decoy system alone, both channels are essential for the circuit's overall functionality, suggesting that that this type of joint control may represent a minimal optimal architecture in different contexts.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper defines a deterministic mass-action model of miRNA regulation during early myogenesis and claims that two independent biosynthesis channels (a decoy system regulated by an exogenous controller and transcription from a distinct genomic locus) are both required: the decoy channel constrains kinetic parameters to narrow ranges for static optimality via competition, while the second locus supplies the nonlinear dynamic response needed for differentiation; competition for miRNA binding is the central control mechanism, and the joint architecture is presented as a minimal optimal design.
Significance. If the model predictions are robust, the work would supply a concrete, quantitative rationale for why dual-channel miRNA control can be necessary for both steady-state precision and rapid transitions, with potential generality to other differentiation circuits. The deterministic formulation permits explicit identification of parameter windows and response nonlinearities, which is a strength when the ranges can be anchored experimentally.
major comments (2)
- [Abstract] Abstract: the central claim that 'both channels are essential for the circuit's overall functionality' and that the decoy system 'constrains kinetic parameters in narrow ranges' rests entirely on internal simulation outcomes; no model equations, parameter values, simulation protocols, or comparison to measured rates are supplied, so the necessity conclusion cannot be evaluated independently of the model's own assumptions.
- [Model construction and results] Model construction and results: the assertions that the channels operate independently and that the identified narrow kinetic windows are biologically required are load-bearing for the 'minimal optimal architecture' conclusion, yet the manuscript provides no external experimental benchmarks or robustness tests against plausible cross-talk or shared machinery that could relax the necessity of joint control.
minor comments (1)
- [Abstract] The abstract would be clearer if it briefly stated the form of the governing equations or the key observables used to define 'static optimality' and 'non-linear response'.
Simulated Author's Rebuttal
We thank the referee for the constructive comments. Below we respond point-by-point to the major comments. As a purely theoretical modeling study, our claims are based on systematic exploration of the deterministic model; we clarify the scope and indicate where revisions can strengthen the presentation.
read point-by-point responses
-
Referee: [Abstract] Abstract: the central claim that 'both channels are essential for the circuit's overall functionality' and that the decoy system 'constrains kinetic parameters in narrow ranges' rests entirely on internal simulation outcomes; no model equations, parameter values, simulation protocols, or comparison to measured rates are supplied, so the necessity conclusion cannot be evaluated independently of the model's own assumptions.
Authors: The abstract provides a concise summary of the findings. The full deterministic mass-action model, including all equations, parameter values (drawn from literature where possible), simulation protocols, and the systematic parameter sweeps demonstrating the narrow windows and necessity of both channels, are presented in the Methods and Results sections. The necessity conclusion is evaluated by comparing the joint architecture against reduced models lacking one channel. We will revise the abstract to explicitly reference the modeling framework and key simulation approach for improved clarity. revision: yes
-
Referee: [Model construction and results] Model construction and results: the assertions that the channels operate independently and that the identified narrow kinetic windows are biologically required are load-bearing for the 'minimal optimal architecture' conclusion, yet the manuscript provides no external experimental benchmarks or robustness tests against plausible cross-talk or shared machinery that could relax the necessity of joint control.
Authors: The model assumes independent channels based on the distinct biological mechanisms (exogenous decoy regulation versus separate genomic transcription locus) reported in the motivating experimental literature. Independence is an input assumption, not a derived claim; the narrow windows emerge from the requirement for effective competition-mediated static control. We conduct extensive robustness checks within the deterministic framework by varying parameters and initial conditions. Explicit modeling of cross-talk or shared machinery is not included, as it would introduce additional unanchored assumptions; the paper presents the joint architecture as a candidate minimal design suggested by the model rather than a proven biological requirement. revision: partial
- Direct comparison to measured experimental rates or provision of new external experimental benchmarks, as this is a theoretical modeling study without new data collection or fitting to specific measurements.
Circularity Check
No circularity: standard mass-action model analyzed on its own terms
full rationale
The paper constructs a deterministic mass-action model from standard biochemical rate equations, then numerically explores its parameter space to identify ranges supporting static optimality via the decoy channel and nonlinear dynamic response via the second locus. The claim that both channels are required for full functionality is an output of that exploration rather than an input presupposed by definition, fitting, or self-citation. No load-bearing step reduces to its own inputs by construction, and the derivation remains self-contained against the model's internal dynamics.
Axiom & Free-Parameter Ledger
free parameters (1)
- kinetic parameters for transcription and binding rates
axioms (2)
- domain assumption The regulatory circuit can be represented by a deterministic system of ordinary differential equations based on mass-action kinetics for miRNA production, binding, and target regulation.
- domain assumption The two miRNA biosynthesis channels operate independently and competition for miRNA binding is the primary mechanism controlling molecular levels.
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel contradicts?
contradictsCONTRADICTS: the theorem conflicts with this paper passage, or marks a claim that would need revision before publication.
optimal static control via the miRNA-decoy system constrains kinetic parameters in narrow ranges where the channels are tightly cross-linked
-
IndisputableMonolith/Foundation/RealityFromDistinction.leanreality_from_one_distinction unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
both channels are essential for the circuit's overall functionality
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
T. R. Cech, J. A. Steitz, The noncoding RNA revolution–trashing old rules to forge new ones, Cell 157 (1) (2014) 77–94
work page 2014
-
[2]
S. M. Hammond, E. Bernstein, D. Beach, G. J. Hannon, An RNA-directed nuclease mediates post-transcriptional gene silencing in Drosophila cells, Nature 404 (6775) (2000) 293–296
work page 2000
-
[3]
D. P. Bartel, MicroRNAs: genomics, biogenesis, mechanism, and function, Cell 116 (2) (2004) 281–297
work page 2004
-
[4]
D. P. Bartel, MicroRNAs: target recognition and regulatory functions, Cell 136 (2) (2009) 215–233
work page 2009
-
[5]
D. Kim, Y . M. Sung, J. Park, S. Kim, J. Kim, J. Park, H. Ha, J. Y . Bae, S. Kim, D. Baek, General rules for functional microRNA targeting, Nature Genetics 48 (12) (2016) 1517–1526
work page 2016
-
[6]
C. P. Ponting, P. L. Oliver, W. Reik, Evolution and functions of long noncoding RNAs, Cell 136 (4) (2009) 629–641
work page 2009
-
[7]
J. L. Rinn, H. Y . Chang, Genome regulation by long noncoding RNAs, Annual Review of Biochemistry 81 (2012) 145–166
work page 2012
- [8]
-
[9]
J. M. Engreitz, N. Ollikainen, M. Guttman, Long non-coding RNAs: spatial amplifiers that control nuclear structure and gene expression, Nature Reviews Molecular Cell Biology 17 (2016) 756–770
work page 2016
-
[10]
V . Siciliano, I. Garzilli, C. Fracassi, S. Criscuolo, S. Ventre, D. di Bernardo, MiRNAs confer phenotypic robustness to gene networks by suppressing biological noise, Nature Communications 4 (2013) 2364
work page 2013
-
[11]
H.-Y . M. Cheng, J. W. Papp, O. Varlamova, H. Dziema, B. Russell, J. P. Curfman, T. Nakazawa, K. Shimizu, H. Okamura, S. Impey, et al., microRNA modulation of circadian-clock period and entrainment, Neuron 54 (5) (2007) 813–829
work page 2007
- [12]
-
[13]
X. Lai, O. Wolkenhauer, J. Vera, Understanding microRNA-mediated gene regulatory networks through mathematical modelling, Nucleic Acids Research 44 (2016) 6019–6035
work page 2016
- [14]
-
[15]
A. Martirosyan, M. Figliuzzi, E. Marinari, A. De Martino, Probing the limits to microRNA-mediated control of gene expression, PLoS Computational Biology 12 (1) (2016) e1004715
work page 2016
-
[16]
A. Martirosyan, A. De Martino, A. Pagnani, E. Marinari, ceRNA crosstalk stabilizes protein expression and a ffects the correlation pattern of interacting proteins, Scientific Reports 7 (2017) 43673
work page 2017
-
[17]
M. Jens, N. Rajewsky, Competition between target sites of regulators shapes post-transcriptional gene regulation, Nature Reviews Genetics 16 (2) (2015) 113–126
work page 2015
-
[18]
L. Salmena, L. Poliseno, Y . Tay, L. Kats, P. P. Pandolfi, A ceRNA hypothesis: The rosetta stone of a hidden RNA language?, Cell 146 (3) (2011) 353–358
work page 2011
-
[19]
M. Figliuzzi, E. Marinari, A. De Martino, MicroRNAs as a selective channel of communication between competing RNAs: a steady-state theory, Biophysical journal 104 (5) (2013) 1203–1213
work page 2013
- [20]
-
[21]
J. Noorbakhsh, A. H. Lang, P. Mehta, Intrinsic noise of microRNA-regulated genes and the ceRNA hypothesis, PLoS One 8 (8) (2013) e72676
work page 2013
-
[22]
M. Figliuzzi, A. De Martino, E. Marinari, RNA-based regulation: dynamics and response to perturbations of competing RNAs, Biophysical journal 107 (4) (2014) 1011–1022
work page 2014
-
[23]
A. D. Bosson, J. R. Zamudio, P. A. Sharp, Endogenous miRNA and target concentrations determine susceptibility to potential ceRNA competition, Molecular cell 56 (3) (2014) 347–359
work page 2014
-
[24]
Y . Yuan, B. Liu, P. Xie, M. Q. Zhang, Y . Li, Z. Xie, X. Wang, Model-guided quantitative analysis of microRNA-mediated regulation on competing endogenous RNAs using a synthetic gene circuit, Proceedings of the National Academy of Sciences 112 (10) (2015) 3158–3163
work page 2015
-
[25]
R. Denzler, S. E. McGeary, A. C. Title, V . Agarwal, D. P. Bartel, M. Stoffel, Impact of MicroRNA levels, Target-Site Complementarity, and Cooperativity on Competing Endogenous RNA-Regulated Gene Expression, Molecular cell 64 (3) (2016) 565–579
work page 2016
-
[26]
L. Poliseno, L. Salmena, J. Zhang, B. Carver, W. J. Haveman, P. P. Pandolfi, A coding-independent function of gene and pseudogene mRNAs regulates tumour biology, Nature 465 (7301) (2010) 1033–1038
work page 2010
-
[27]
J. Wang, X. Liu, H. Wu, P. Ni, Z. Gu, Y . Qiao, N. Chen, F. Sun, Q. Fan, CREB up-regulates long non-coding RNA, HULC expression through interaction with microRNA-372 in liver cancer, Nucleic Acids Research 38 (16) (2010) 5366–5383
work page 2010
-
[28]
F. A. Karreth, M. Reschke, A. Ruocco, C. Ng, B. Chapuy, V . L ´eopold, M. Sjoberg, T. M. Keane, A. Verma, U. Ala, et al., The BRAF pseudogene functions as a competitive endogenous RNA and induces lymphoma in vivo, Cell 161 (2) (2015) 319–332
work page 2015
- [29]
-
[30]
M. V . Neguembor, M. Jothi, D. Gabellini, Long noncoding RNAs, emerging players in muscle di fferentiation and disease, Skeletal muscle 4 (1) (2014) 8
work page 2014
-
[31]
I. Legnini, M. Morlando, A. Mangiavacchi, A. Fatica, I. Bozzoni, A feedforward regulatory loop between HuR and the long noncoding RNA linc-MD1 controls early phases of myogenesis, Molecular cell 53 (3) (2014) 506–514
work page 2014
-
[32]
H. Shen, A. S. McElhinny, Y . Cao, P. Gao, J. Liu, R. Bronson, J. D. Gri ffin, L. Wu, The Notch coactivator, MAML1, functions as a novel coactivator for MEF2C-mediated transcription and is required for normal myogenesis, Genes & development 20 (6) (2006) 675–688
work page 2006
-
[33]
M. A. Valencia-Sanchez, J. Liu, G. J. Hannon, R. Parker, Control of translation and mRNA degradation by miRNAs and siRNAs, Genes & development 20 (5) (2006) 515–524
work page 2006
-
[34]
X. Wang, Y . Li, X. Xu, Y .-h. Wang, Toward a system-level understanding of microRNA pathway via mathematical modeling, Biosystems 100 (1) (2010) 31–38. 16
work page 2010
- [35]
-
[36]
Alon, An introduction to systems biology: design principles of biological circuits, CRC press, 2006
U. Alon, An introduction to systems biology: design principles of biological circuits, CRC press, 2006
work page 2006
-
[37]
Palsson, Systems biology: properties of reconstructed networks
B. Palsson, Systems biology: properties of reconstructed networks. 2006, Cambridge Univ Press
work page 2006
-
[38]
A. Martirosyan, M. Marsili, A. De Martino, Translating ceRNA susceptibilities into correlation functions, bioRxiv preprint [http://www.biorxiv.org/content/early/2017/01/25/102988]. 17
work page 2017
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.