Dimensionality reduction of neuronal degeneracy reveals two interfering physiological mechanisms
Pith reviewed 2026-05-24 01:20 UTC · model grok-4.3
The pith
Dimensionality reduction of ion channel conductances reveals two main sources of variability driven by activity regulation feedback.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
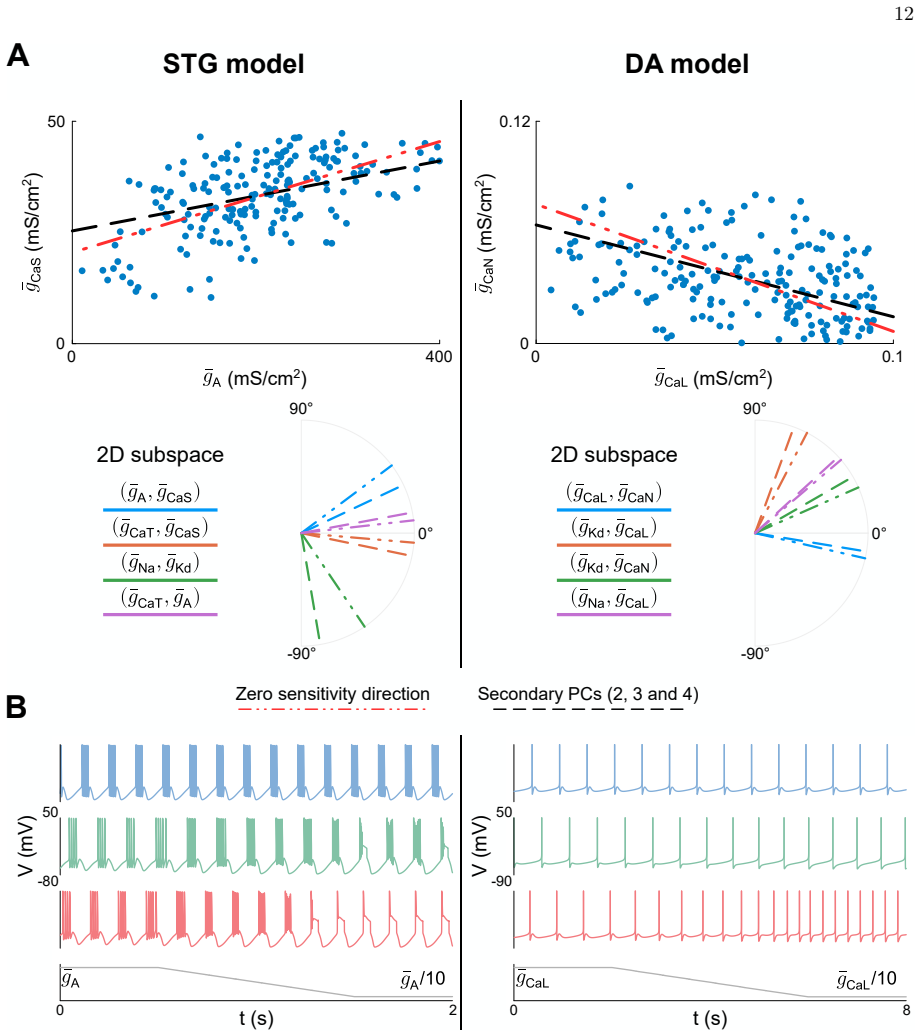
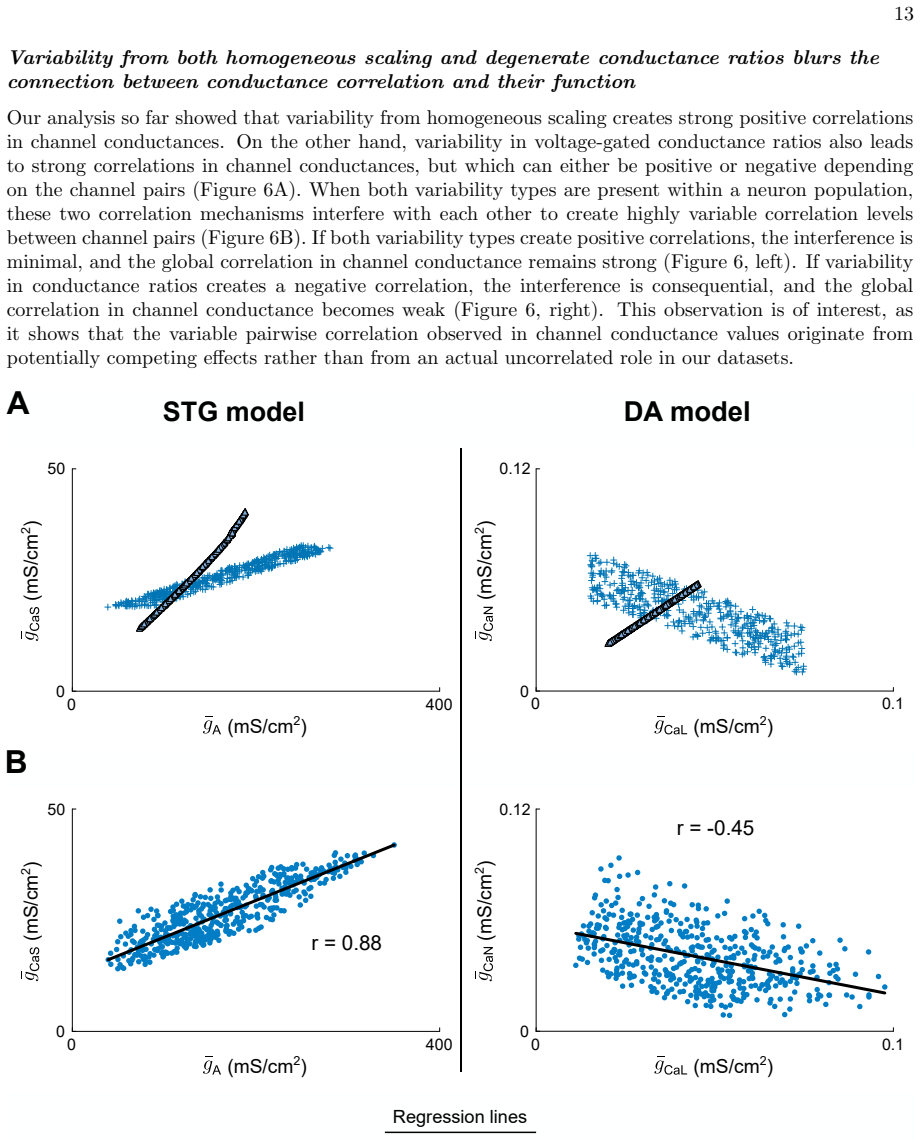
Using detailed conductance-based modeling to explore the origin of stable neuronal function from variable channel composition, dimensionality reduction uncovers two principal dimensions in the channel conductance space that capture most of the variance of the observed variability. Those two dimensions correspond to two physiologically relevant sources of variability that can be explained by feedback mechanisms underlying regulation of neuronal activity, providing quantitative insights into how channel composition links to neuronal electrophysiological activity. These insights allowed the design of a model-independent, reliable neuromodulation rule for variable neuronal populations.
What carries the argument
Two principal components of the ion-channel conductance space that align with distinct activity-dependent feedback regulation loops.
If this is right
- Most variability in channel expression arises from two interfering physiological mechanisms rather than many independent sources.
- Channel composition determines electrophysiological activity through these two low-dimensional directions.
- A single neuromodulation rule can be applied reliably to neuronal populations that differ widely in their exact channel numbers.
- Changes made along either dimension produce predictable shifts in firing behavior that can be anticipated without full model re-simulation.
Where Pith is reading between the lines
- The same two-dimensional structure may appear in other excitable cells whose function must remain stable despite conductance variation.
- Targeted experiments that perturb one feedback loop while measuring the resulting conductance covariance could confirm or refute the mapping to the two principal components.
- The reduction offers a practical route to simplify large-scale network models by replacing many independent channel parameters with two effective variables.
Load-bearing premise
Variability generated inside the conductance-based models accurately represents biological differences in ion-channel expression, and the two principal components map directly onto separate physiological feedback mechanisms.
What would settle it
Record channel conductances across a population of real neurons that share the same electrophysiological phenotype; if the measured spread does not lie mostly along the two identified dimensions, or if a neuromodulation rule based on those dimensions fails to stabilize activity, the central claim is refuted.
Figures






read the original abstract
Neuronal systems maintain stable functions despite large variability in their physiological components. Ion channel expression, in particular, is highly variable in neurons exhibiting similar electrophysiological phenotypes, which poses questions regarding how specific ion channel subsets reliably shape neuron intrinsic properties. Here, we use detailed conductance-based modeling to explore the origin of stable neuronal function from variable channel composition. Using dimensionality reduction, we uncover two principal dimensions in the channel conductance space that capture most of the variance of the observed variability. Those two dimensions correspond to two physiologically relevant sources of variability that can be explained by feedback mechanisms underlying regulation of neuronal activity, providing quantitative insights into how channel composition links to neuronal electrophysiological activity. These insights allowed us to understand and design a model-independent, reliable neuromodulation rule for variable neuronal populations.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript uses detailed conductance-based models to generate populations of neurons with variable ion channel conductances but stable electrophysiological phenotypes. Dimensionality reduction is applied to the conductance space to identify two principal dimensions capturing most observed variance; these are interpreted as corresponding to two distinct physiological feedback mechanisms regulating neuronal activity. The resulting framework is used to derive a model-independent neuromodulation rule applicable to variable neuronal populations.
Significance. If the mapping from the reduced dimensions to specific feedback mechanisms can be shown to hold independently of modeling choices, the work would provide a quantitative approach to understanding neuronal degeneracy and could inform more robust neuromodulation strategies that accommodate biological variability in channel expression.
major comments (2)
- [Abstract] Abstract: the central claim that the two principal dimensions 'correspond to two physiologically relevant sources of variability that can be explained by feedback mechanisms' is presented as an interpretive step without any described independent validation, error quantification, or explicit mapping procedure; this interpretive link is load-bearing for both the physiological interpretation and the downstream model-independent neuromodulation rule.
- [Abstract] Abstract: the assertion that the approach yields a 'model-independent, reliable neuromodulation rule' requires demonstration that perturbations along the identified axes reproduce the dynamics of the hypothesized feedback loops rather than reflecting the particular choice of model equations or sampling distribution; no such check is described.
minor comments (2)
- The abstract does not specify the exact dimensionality reduction technique employed (e.g., PCA) or the fraction of variance captured by the two dimensions; these quantitative details should appear in the opening summary.
- No information is given on the number of model instances, the ranges or distributions used for conductance sampling, or any cross-validation of the reduced space; these would strengthen assessment of robustness.
Simulated Author's Rebuttal
We thank the referee for their constructive comments, which highlight important aspects of validation and generality in our work. We address each major comment below and indicate where revisions will be made to strengthen the manuscript.
read point-by-point responses
-
Referee: [Abstract] Abstract: the central claim that the two principal dimensions 'correspond to two physiologically relevant sources of variability that can be explained by feedback mechanisms' is presented as an interpretive step without any described independent validation, error quantification, or explicit mapping procedure; this interpretive link is load-bearing for both the physiological interpretation and the downstream model-independent neuromodulation rule.
Authors: The mapping from the two principal components to feedback mechanisms is based on the alignment between the directions of maximal variance in the sampled conductance space and the compensatory adjustments expected from established activity-dependent regulatory processes (e.g., calcium-dependent or firing-rate homeostatic rules). This alignment was quantified by projecting the conductance vectors of phenotype-matched models onto the PC axes and verifying that the resulting electrophysiological changes remain within the target phenotype bounds. We acknowledge that the manuscript does not include an independent validation step (such as cross-model testing or direct comparison to experimental channel-expression data) or explicit error bounds on the mapping. We will revise the Methods and Results sections to provide a detailed, step-by-step description of the mapping procedure together with quantitative metrics (e.g., explained variance per axis and correlation with known feedback targets). revision: yes
-
Referee: [Abstract] Abstract: the assertion that the approach yields a 'model-independent, reliable neuromodulation rule' requires demonstration that perturbations along the identified axes reproduce the dynamics of the hypothesized feedback loops rather than reflecting the particular choice of model equations or sampling distribution; no such check is described.
Authors: The neuromodulation rule is constructed by treating the two principal axes as the directions along which conductances can be co-varied while preserving the target electrophysiological phenotype; this is verified by applying controlled perturbations along each axis to held-out model instances and confirming that spike-frequency and other core properties remain stable. We agree that the current presentation does not explicitly test whether the same axes emerge under alternative model formulations or different sampling distributions of channel conductances. We will add a supplementary analysis that applies the identical dimensionality-reduction pipeline to (i) a second conductance-based model with different channel kinetics and (ii) a re-sampled population drawn from a broader prior, thereby quantifying the degree of axis stability across modeling choices. revision: yes
Circularity Check
No significant circularity; derivation is self-contained.
full rationale
The paper generates variability via parameter sampling in conductance-based models, applies dimensionality reduction to identify principal components capturing variance in that sampled space, and then interprets the resulting axes as corresponding to feedback mechanisms. This interpretation is presented as a post-analysis insight rather than a definitional or fitted reduction to the inputs. No quoted equations or self-citations reduce the central claim (PCA dimensions indexing physiological sources, enabling neuromodulation rule) to tautology by construction. The analysis remains independent of the specific sampling distribution once the reduction is performed, satisfying the criteria for a non-circular finding.
Axiom & Free-Parameter Ledger
axioms (1)
- domain assumption Feedback mechanisms underlie regulation of neuronal activity and can be recovered from principal components of conductance variability.
Reference graph
Works this paper leans on
-
[1]
Achard, P. and De Schutter, E. (2006). Complex parameter landscape for a complex neuron model. PLoS computational biology , 2(7):e94
work page 2006
-
[2]
Alonso, L. M. and Marder, E. (2019). Visualization of currents in neural models with similar behavior and different conductance densities. Elife , 8:e42722
work page 2019
-
[3]
Amendola, J., Woodhouse, A., Martin-Eauclaire, M.-F., and Goaillard, J.-M. (2012). Ca2+/camp-sensitive covariation of ia and ih voltage dependences tunes rebound firing in dopaminergic neurons. Journal of Neuroscience , 32(6):2166--2181
work page 2012
-
[4]
Bezanson, J., Edelman, A., Karpinski, S., and Shah, V. B. (2017). Julia: A fresh approach to numerical computing. SIAM review , 59(1):65--98
work page 2017
-
[5]
Drion, G., Franci, A., Dethier, J., and Sepulchre, R. (2015a). Dynamic input conductances shape neuronal spiking. eneuro , 2(1)
-
[6]
Drion, G., O’Leary, T., and Marder, E. (2015b). Ion channel degeneracy enables robust and tunable neuronal firing rates. Proceedings of the National Academy of Sciences , 112(38):E5361--E5370
-
[7]
Franci, A., Drion, G., and Sepulchre, R. (2018). Robust and tunable bursting requires slow positive feedback. Journal of neurophysiology , 119(3):1222--1234
work page 2018
-
[8]
Goaillard, J.-M. and Marder, E. (2021). Ion channel degeneracy, variability, and covariation in neuron and circuit resilience. Annual review of neuroscience , 44:335--357
work page 2021
-
[9]
Grashow, R., Brookings, T., and Marder, E. (2009). Reliable neuromodulation from circuits with variable underlying structure. Proceedings of the National Academy of Sciences , 106(28):11742--11746
work page 2009
-
[10]
A., Hampton, D., and Marder, E
Haley, J. A., Hampton, D., and Marder, E. (2018). Two central pattern generators from the crab, cancer borealis, respond robustly and differentially to extreme extracellular ph. Elife , 7:e41877
work page 2018
-
[11]
Iacobas, D. A., Iacobas, S., Lee, P. R., Cohen, J. E., and Fields, R. D. (2019). Coordinated activity of transcriptional networks responding to the pattern of action potential firing in neurons. Genes , 10(10):754
work page 2019
-
[12]
Khorkova, O. and Golowasch, J. (2007). Neuromodulators, not activity, control coordinated expression of ionic currents. Journal of Neuroscience , 27(32):8709--8718
work page 2007
-
[13]
H., Shin, M., Kelleher, K., Kolkman, K
Kodama, T., Gittis, A. H., Shin, M., Kelleher, K., Kolkman, K. E., McElvain, L., Lam, M., and Du Lac, S. (2020). Graded coexpression of ion channel, neurofilament, and synaptic genes in fast-spiking vestibular nucleus neurons. Journal of Neuroscience , 40(3):496--508
work page 2020
-
[14]
Kramer, R. H. and Levitan, I. B. (1990). Activity-dependent neuromodulation in aplysia neuron r15: intracellular calcium antagonizes neurotransmitter responses mediated by camp. Journal of neurophysiology , 63(5):1075--1088
work page 1990
-
[15]
Liss, B., Franz, O., Sewing, S., Bruns, R., Neuhoff, H., and Roeper, J. (2001). Tuning pacemaker frequency of individual dopaminergic neurons by kv4. 3l and kchip3. 1 transcription. The EMBO journal , 20(20):5715--5724
work page 2001
-
[16]
Liu, Z., Golowasch, J., Marder, E., and Abbott, L. (1998). A model neuron with activity-dependent conductances regulated by multiple calcium sensors. Journal of Neuroscience , 18(7):2309--2320
work page 1998
-
[17]
Marder, E. (2012). Neuromodulation of neuronal circuits: back to the future. Neuron , 76(1):1--11
work page 2012
-
[18]
Marder, E. and Bucher, D. (2007). Understanding circuit dynamics using the stomatogastric nervous system of lobsters and crabs. Annu. Rev. Physiol. , 69:291--316
work page 2007
-
[19]
Marder, E. and Goaillard, J.-M. (2006). Variability, compensation and homeostasis in neuron and network function. Nature Reviews Neuroscience , 7(7):563--574
work page 2006
-
[20]
Marder, E., O'Leary, T., and Shruti, S. (2014). Neuromodulation of circuits with variable parameters: single neurons and small circuits reveal principles of state-dependent and robust neuromodulation. Annual review of neuroscience , 37:329--346
work page 2014
-
[21]
Nadim, F. and Bucher, D. (2014). Neuromodulation of neurons and synapses. Current opinion in neurobiology , 29:48--56
work page 2014
-
[22]
O'Leary, T., Williams, A. H., Caplan, J. S., and Marder, E. (2013). Correlations in ion channel expression emerge from homeostatic tuning rules. Proceedings of the National Academy of Sciences , 110(28):E2645--E2654
work page 2013
-
[23]
O’Leary, T., Williams, A. H., Franci, A., and Marder, E. (2014). Cell types, network homeostasis, and pathological compensation from a biologically plausible ion channel expression model. Neuron , 82(4):809--821
work page 2014
-
[24]
O’Leary, T. and Wyllie, D. J. (2011). Neuronal homeostasis: time for a change? The Journal of physiology , 589(20):4811--4826
work page 2011
-
[25]
Prinz, A. A., Bucher, D., and Marder, E. (2004). Similar network activity from disparate circuit parameters. Nature neuroscience , 7(12):1345--1352
work page 2004
-
[26]
Qian, K., Yu, N., Tucker, K. R., Levitan, E. S., and Canavier, C. C. (2014). Mathematical analysis of depolarization block mediated by slow inactivation of fast sodium channels in midbrain dopamine neurons. Journal of neurophysiology , 112(11):2779--2790
work page 2014
-
[27]
Raymond, J. L., Baxter, D. A., Buonomano, D. V., and Byrne, J. H. (1992). A learning rule based on empirically-derived activity-dependent neuromodulation supports operant conditioning in a small network. Neural Networks , 5(5):789--803
work page 1992
-
[28]
Rinberg, A., Taylor, A. L., and Marder, E. (2013). The effects of temperature on the stability of a neuronal oscillator. PLoS computational biology , 9(1):e1002857
work page 2013
-
[29]
Santin, J. M. and Schulz, D. J. (2019). Membrane voltage is a direct feedback signal that influences correlated ion channel expression in neurons. Current Biology , 29(10):1683--1688
work page 2019
-
[30]
Schultz, W. (2007). Multiple dopamine functions at different time courses. Annu. Rev. Neurosci. , 30:259--288
work page 2007
-
[31]
Schulz, D. J., Baines, R. A., Hempel, C. M., Li, L., Liss, B., and Misonou, H. (2006a). Cellular excitability and the regulation of functional neuronal identity: from gene expression to neuromodulation. Journal of Neuroscience , 26(41):10362--10367
-
[32]
J., Goaillard, J.-M., and Marder, E
Schulz, D. J., Goaillard, J.-M., and Marder, E. (2006b). Variable channel expression in identified single and electrically coupled neurons in different animals. Nature neuroscience , 9(3):356--362
-
[33]
J., Goaillard, J.-M., and Marder, E
Schulz, D. J., Goaillard, J.-M., and Marder, E. E. (2007). Quantitative expression profiling of identified neurons reveals cell-specific constraints on highly variable levels of gene expression. Proceedings of the National Academy of Sciences , 104(32):13187--13191
work page 2007
-
[34]
Swensen, A. M. and Bean, B. P. (2005). Robustness of burst firing in dissociated purkinje neurons with acute or long-term reductions in sodium conductance. Journal of Neuroscience , 25(14):3509--3520
work page 2005
-
[35]
Tapia, M., Baudot, P., Formisano-Tr \'e ziny, C., Dufour, M. A., Temporal, S., Lasserre, M., Marqu \`e ze-Pouey, B., Gabert, J., Kobayashi, K., and Goaillard, J.-M. (2018). Neurotransmitter identity and electrophysiological phenotype are genetically coupled in midbrain dopaminergic neurons. Scientific reports , 8(1):13637
work page 2018
-
[36]
L., Goaillard, J.-M., and Marder, E
Taylor, A. L., Goaillard, J.-M., and Marder, E. (2009). How multiple conductances determine electrophysiological properties in a multicompartment model. Journal of Neuroscience , 29(17):5573--5586
work page 2009
-
[37]
Temporal, S., Desai, M., Khorkova, O., Varghese, G., Dai, A., Schulz, D. J., and Golowasch, J. (2012). Neuromodulation independently determines correlated channel expression and conductance levels in motor neurons of the stomatogastric ganglion. Journal of neurophysiology , 107(2):718--727
work page 2012
-
[38]
Tobin, A.-E., Cruz-Berm \'u dez, N. D., Marder, E., and Schulz, D. J. (2009). Correlations in ion channel mrna in rhythmically active neurons. PloS one , 4(8):e6742
work page 2009
-
[39]
Walters, E. T. and Byrne, J. H. (1984). Activity-dependent neuromodulation: A mechanism for associative plasticity. Neuronal Growth and Plasticity , 6:219
work page 1984
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.