Building and maintaining a System of Intracellular Compartments
Pith reviewed 2026-05-22 10:57 UTC · model grok-4.3
The pith
The vesicular transport and cisternal progression models of the Golgi are two phases of one nonequilibrium process based on fusion-fission cycles.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
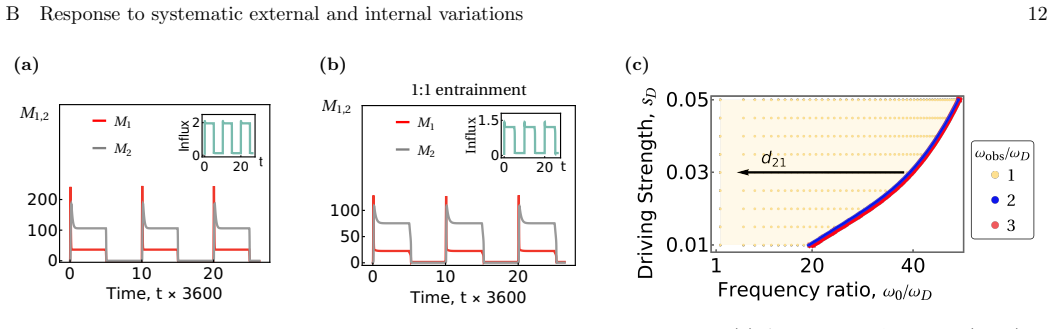
Within a stochastic framework of mechanochemical fusion-fission cycles that violate detailed balance, the system exhibits distinct robust regimes ranging from fixed points to limit cycles with definite phase relations between cisternae. These regimes map onto diverse phenotypes including stable cisternae, cell-cycle-dependent dissolution and reassembly, and cisternal progression. The analysis shows that the vesicular transport and cisternal progression models are two phases of the same underlying nonequilibrium process, with cisternal size homeostasis achieved by an embedded control system driven by size-dependent fusion-fission kernels.
What carries the argument
The stochastic framework of mechanochemical fusion-fission cycles that violate detailed balance, which generates robust dynamical regimes and embeds size control through size-dependent kernels.
If this is right
- The two competing models of Golgi organization emerge as different phases of the same process.
- Size control of cisternae arises naturally from size-dependent fusion and fission rates.
- Dynamical regimes correspond to stable structures, periodic cycling, and progressive maturation.
- Modulating the interplay between glycosylation enzymes and membrane dynamics can control cisternal number and chemical identity.
- The framework makes definite predictions for the system's response to perturbations that can be tested experimentally.
Where Pith is reading between the lines
- This approach could extend to understanding the organization of other membrane-bound organelles in the cell.
- Similar nonequilibrium mechanisms might govern the maintenance of compartment identity in endocytic pathways.
- Engineered cells could use modulated fusion-fission dynamics to achieve desired organelle configurations.
- Connections to broader nonequilibrium physics in biological self-assembly suggest testable links to other cellular structures.
Load-bearing premise
A stochastic framework of mechanochemical fusion-fission cycles that violate detailed balance is sufficient to generate the observed robust regimes and to embed size control via size-dependent kernels.
What would settle it
An experiment that perturbs fusion-fission rates and fails to observe the predicted switching between stable cisternae, periodic reassembly, and cisternal progression, or that shows no size-dependent control, would challenge the central claim.
Figures






read the original abstract
Organelle patterning and its heritability remain central mysteries in cell biology, highlighting the fundamental tension between genetic inheritance and self-assembly. Here, we explore the nonequilibrium assembly and emdedded size control of the Golgi cisternae and endosomes, amid a continuous flux of membrane traffic, within a stochastic framework of mechanochemical fusion-fission cycles that violate detailed balance. Using a dynamical systems approach, we identify distinct, robust regimes, ranging from fixed points to limit cycles with definite phase relations between cisternae. We identify these dynamical regimes with diverse phenotypes, from stable cisternae to periodic, cell-cycle-dependent dissolution/reassembly of cisternae to cisternal progression. We analyse its dynamic response to systematic perturbations or driving protocols and make definite predictions that may be tested experimentally. Our analysis reveals that the two competing models of Golgi organization - vesicular transport and cisternal progression - are, in fact, two phases of the same underlying nonequilibrium process. We see that cisternal size homeostasis is brought about by a size-dependent embedded control system driven by fusion-fission kernels. Finally, our framework offers a strategy for controlling cisternal number and chemical identity by modulating the interplay between glycosylation enzymes and membrane fission-fusion dynamics.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript develops a stochastic dynamical-systems model of Golgi cisternae and endosomes driven by mechanochemical fusion-fission cycles that violate detailed balance. It identifies robust regimes (fixed points and limit cycles with definite phase relations) and maps them onto biological phenotypes ranging from stable cisternae to periodic cell-cycle-dependent reassembly and cisternal progression. The central claim is that vesicular transport and cisternal progression are two phases of the same nonequilibrium process, with cisternal size homeostasis arising from an embedded size-dependent control system encoded in the fusion-fission kernels. The work also examines dynamic responses to perturbations and offers testable experimental predictions.
Significance. If the dynamical analysis is correct, the paper supplies a unifying nonequilibrium framework that reconciles two long-standing competing models of Golgi organization within a single stochastic process. It demonstrates how size control and chemical identity can emerge from mechanochemical cycles without external templating, and it supplies concrete, falsifiable predictions. These strengths would make the work a notable contribution at the interface of soft-matter physics and cell biology.
major comments (2)
- [Abstract and §4] Abstract and §4 (or equivalent dynamical-systems section): the claim that vesicular transport and cisternal progression are phases of one process requires an explicit bifurcation or continuous phase boundary in parameter space that connects the fixed-point regime to the limit-cycle regime under variation of a single, biologically interpretable parameter (e.g., fission rate, glycosylation enzyme density, or membrane tension). The abstract identifies distinct regimes but does not indicate whether they are connected by a continuous path rather than by disconnected parameter choices.
- [Model definition (likely §2)] Model definition (likely §2): the size-dependent kernels are stated to embed size control, yet it is unclear whether these kernels are derived from the mechanochemical rules or introduced as phenomenological functions. If the latter, the 'embedded control system' claim risks being circular; the manuscript should show that the kernels follow from the underlying fusion-fission energetics without additional fitting.
minor comments (2)
- [Throughout] Notation for the fusion-fission rate kernels should be defined once, with explicit statement of how detailed balance is violated (e.g., via an energy-consuming term).
- [Figures] Phase diagrams or bifurcation plots (if present) should label the control parameter on the axis and indicate its biological counterpart.
Simulated Author's Rebuttal
We thank the referee for the positive and constructive report. The assessment of the work's significance at the interface of soft-matter physics and cell biology is appreciated. We address each of the major comments below and will make revisions to the manuscript as indicated.
read point-by-point responses
-
Referee: [Abstract and §4] Abstract and §4 (or equivalent dynamical-systems section): the claim that vesicular transport and cisternal progression are phases of one process requires an explicit bifurcation or continuous phase boundary in parameter space that connects the fixed-point regime to the limit-cycle regime under variation of a single, biologically interpretable parameter (e.g., fission rate, glycosylation enzyme density, or membrane tension). The abstract identifies distinct regimes but does not indicate whether they are connected by a continuous path rather than by disconnected parameter choices.
Authors: We agree that an explicit demonstration of a continuous connection between the regimes would strengthen the central claim that they represent phases of the same process. In the revised manuscript, we will include a bifurcation analysis in the dynamical systems section (§4), varying a single parameter such as the fission rate. This will show a continuous transition, for example via a Hopf bifurcation, from the fixed-point regime associated with stable cisternae and vesicular transport to the limit-cycle regime corresponding to cisternal progression. We will update the abstract to reflect this connection. revision: yes
-
Referee: [Model definition (likely §2)] Model definition (likely §2): the size-dependent kernels are stated to embed size control, yet it is unclear whether these kernels are derived from the mechanochemical rules or introduced as phenomenological functions. If the latter, the 'embedded control system' claim risks being circular; the manuscript should show that the kernels follow from the underlying fusion-fission energetics without additional fitting.
Authors: The size-dependent kernels are motivated by and derived from the mechanochemical fusion-fission rules, incorporating the effects of cisternal size on membrane tension and the resulting fusion and fission probabilities. This is outlined in the model definition. To prevent any perception of circularity, we will revise the manuscript to provide a more detailed step-by-step derivation showing how the kernels arise directly from the underlying energetics, without introducing additional fitting parameters beyond the basic physical assumptions of the model. revision: partial
Circularity Check
Size homeostasis via size-dependent kernels risks being self-definitional
specific steps
-
self definitional
[Abstract]
"We see that cisternal size homeostasis is brought about by a size-dependent embedded control system driven by fusion-fission kernels."
Homeostasis is claimed to result from the size-dependent kernels, yet the kernels are introduced as size-dependent precisely to produce this control; the outcome is then equivalent to the modeling choice rather than derived from the underlying stochastic fusion-fission dynamics that violate detailed balance.
full rationale
The abstract asserts that size homeostasis arises from a size-dependent embedded control system driven by fusion-fission kernels within the stochastic nonequilibrium framework. Without the full equations, this risks reducing to an input assumption rather than an independent derivation from the mechanochemical cycles. The unification of vesicular transport and cisternal progression as phases of one process is presented as a dynamical-systems result, but the provided text does not exhibit an explicit bifurcation or continuous transition under a single tunable biological parameter; separate regimes could be obtained by disconnected parameter choices. No self-citation load-bearing or uniqueness theorem is visible in the abstract. The central claims retain independent modeling content, so circularity is only partial and moderate. The derivation chain appears self-contained against external benchmarks once the kernels are specified.
Axiom & Free-Parameter Ledger
free parameters (1)
- fusion-fission kernels
axioms (1)
- domain assumption Mechanochemical fusion-fission cycles violate detailed balance
invented entities (1)
-
embedded size control system
no independent evidence
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
K_fus = a + b M^α/(C1+M^α), K_fis = d M^β/(C2+M^β) ... size-dependent negative feedback control via fusion-fission kernels
-
IndisputableMonolith/Foundation/RealityFromDistinction.leanreality_from_one_distinction unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
two competing models ... are, in fact, two phases of the same underlying nonequilibrium process
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
Solve the differential equations Eqs.(S68),(S69) in(𝑀 1, 𝑀2)(Eqs.(12),(13) in the main text) for given values of parameters and initial conditions
-
[2]
Choose, say, five time points𝜏 𝑎 < 𝜏 𝑏 < 𝜏 𝑐 < 𝜏 𝑑 < 𝜏 𝑒, these time points are taken to be large enough numbers to make sure that the system has arrived at a steady-state. We compute the following quantities :|𝑀 1 (𝜏𝑎) − 𝑀1 (𝜏𝑐)|,|𝑀 1 (𝜏𝑏) −𝑀 1 (𝜏𝑑)|,|𝑀 2 (𝜏𝑎) −𝑀 2 (𝜏𝑐)|,|𝑀 2 (𝜏𝑐) −𝑀 2 (𝜏𝑑)|, 𝑀 1 (𝑡=𝜏 𝑒), 𝑀2 (𝑡=𝜏 𝑒)
-
[3]
The steady-state is a fixed point solution if 0<|𝑀 1 (𝜏𝑎) −𝑀 1 (𝜏𝑐)|< 𝛿 1, 0<|𝑀 1 (𝜏𝑏) −𝑀 1 (𝜏𝑑)|< 𝛿 1, 0<|𝑀 2 (𝜏𝑎) − 𝑀2 (𝜏𝑐)|< 𝛿 1, 0<|𝑀 2 (𝜏𝑏) −𝑀 2 (𝜏𝑑)|< 𝛿 1,𝑀 1 (𝑡=𝜏 𝑒)< 𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ and𝑀 2 (𝑡=𝜏 𝑒)< 𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ ,𝛿 1 is the tolerance assumed and𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ =300 is the threshold size assumed for the stable state
-
[4]
The steady-state is a limit cycle solution if𝛿 1 ≤ |𝑀 1 (𝜏𝑎) −𝑀 1 (𝜏𝑐)|< 𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ ,𝛿 1 ≤ |𝑀 1 (𝜏𝑏) −𝑀 1 (𝜏𝑑)|< 𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ , 𝛿1 ≤ |𝑀 2 (𝜏𝑎) −𝑀 2 (𝜏𝑐)|< 𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ ,𝛿 1 ≤ |𝑀 2 (𝜏𝑏) −𝑀 2 (𝜏𝑑)|< 𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ ,𝑀 1 (𝑡=𝜏 𝑒)< 𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ and𝑀 2 (𝑡=𝜏 𝑒)< 𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ
-
[5]
S5.5 Arriving at the phase diagram and phase boundaries for the two-cisternae system 47 No
The system is unbounded if𝑀 1 (𝑡=𝜏 𝑒) ≥𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ or𝑀 2 (𝑡=𝜏 𝑒) ≥𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ . S5.5 Arriving at the phase diagram and phase boundaries for the two-cisternae system 47 No. of distinct roots Algebraic capacity 0 1 2 3 4 5 0 10 20 30 Influx rate, v Root count, r N rN =18 Figure S14:Structural stability of two-cisternae dynamical system. The structural stabilit...
-
[6]
While the numerical scheme presented above is adequate for the present purposes, it is subject to the following limitations – (i) it would not detect limit cycle solutions with a radius𝑟 𝜏, such that𝑟 𝜏 < 𝛿 1 or𝑟 𝜏 > 𝑀 𝑡 ℎ – and would assume them to be fixed point solutions. (ii) Fixed point solutions that reach the asymptotic value at a time much larger ...
-
[7]
Compute the Gr¨ obner basis from the system of equations/vector fields (e.g.𝑥 3 +𝑏 𝑥+𝑐for cusp singularity [47], where x is the state variable and b and c are constant parameters). We have usedMathematicafor this (which uses Buchberger algorithm and DegreeReverseLexicographic order, see [86] for details)
-
[8]
Extract the leading monomials from each polynomial in the Gr¨ obner basis (e.g.,{𝑥 3})
-
[9]
Generate all monomials up to a chosen maximum polynomial Degree:{1, 𝑥, 𝑥 2, 𝑥3, 𝑥4, 𝑥5, . . .}
-
[10]
Reject all monomials divisible by any leading monomial (e.g., reject𝑥 3, 𝑥4, 𝑥5, . . .)
-
[11]
The surviving monomials form the standard monomial basis (e.g.,{1, 𝑥, 𝑥 2})
-
[12]
Algebraic capacity = dimension of standard monomial basis (e.g., capacity = 3)
-
[13]
Choose specific numerical values for the parameters and numerically solve the system to count the number of distinct complex roots (geometric roots)
-
[14]
Compare: if geometric roots = algebraic capacity, the system is structurally stable. 48
-
[15]
The above steps for the 2-cisternae system are tabulated in Table S4
If geometric roots<algebraic capacity, the system is structurally unstable. The above steps for the 2-cisternae system are tabulated in Table S4. The number of distinct complex roots and the algebraic capacity are plotted in Fig. S14. Vector field ( 𝑣 𝑎1 + 𝑀2 1 𝐶11 +𝑀 2 1 ! − 𝑑1 𝑀1 𝐶12 +𝑀 1 − 𝑑12 𝑀1 𝐶12 +𝑀 1 𝑎2 + 𝑀2 2 𝐶21 +𝑀 2 2 ! + 𝑑21 𝑀2 𝐶22 +𝑀 2 𝑎1 + 𝑀...
work page 2000
-
[16]
Parameter values,𝑣=2, 𝐶11 =100,𝐶 22 =20,𝐶 12 =20,𝑑 12 =2,𝑑 21 =0,𝑑 1 =1,𝑑 2 =1.5,𝑎 1 =𝑎 2 =0.05
+ ∑︁ 𝑖 𝜏𝑖 𝑆 𝑁 (𝐸 1 𝑖 , 𝐸2 𝑖 , 𝑥1 𝑖 , 𝑥2 𝑖 ) 𝑖∈SN bifurcations encompassed by the trajectory (S105) S8.2 De novo assembly of two cisternae 55 (a) M1 M2 0 500 1000 1500 0 50 100 150 200 Time, t M1,2 τSN τSN+τf (b) (c) M1 M2 0 500 1000 15000 20 40 Time, t M2 τSN τSN+τ0 (d) Figure S18:Formation time for two cisternae.(a) Cisternae formation time for fixed poi...
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.