Universal principles of cell population growth follow from local contact inhibition
Pith reviewed 2026-05-24 13:22 UTC · model grok-4.3
The pith
Local contact inhibition produces five classical cell population growth laws from one microscopic model.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
The connection between microscopic assumptions and expected contact inhibition leads to five classical tumor growth laws: exponential, radial growth, fractal growth, generalized logistic, and Gompertzian growth. All five can be seen as manifestations of a single microscopic model. Agent-based simulations substantiate the theory and explain differences in growth curves from experimental in vitro cancer cell population data.
What carries the argument
Local contact inhibition, the rule that cell proliferation is suppressed by physical contact with neighboring cells, which under differing auxiliary assumptions on mixing or space produces each of the five macroscopic growth laws.
If this is right
- Observed differences in experimental growth curves can be attributed to how contact inhibition interacts with population mixing without additional parameters.
- Many separate mean-field laws for cancer or microbial growth become connected through the same contact inhibition mechanism.
- Quantitative predictions of cell growth shift when contact inhibition is considered together with assumptions about spatial structure.
- Agent-based models under this single rule reproduce the macroscopic behaviors seen across the five laws.
Where Pith is reading between the lines
- Manipulating cell adhesion in experiments could switch a population between different growth laws in a predictable way.
- The same contact rule may describe growth in bacterial colonies or tissue cultures when mixing conditions are controlled.
- The framework suggests testing whether altering local density thresholds changes which macroscopic law best fits the data.
Load-bearing premise
That local contact inhibition combined with standard auxiliary assumptions such as well-mixed conditions produces the exact functional forms of each of the five growth laws without extra fitted parameters.
What would settle it
Observation of cell population growth whose curve matches none of the five laws despite clear local contact inhibition and matching auxiliary conditions on mixing or space.
Figures



read the original abstract
Cancer cell populations often exhibit remarkably similar growth laws despite their heterogeneity. Explanations of universal cell population growth remain partly unresolved to this day. Here, we present a growth-law unification by investigating the connection between microscopic assumptions and the expected contact inhibition, which leads to five classical tumor growth laws: exponential, radial growth, fractal growth, generalized logistic, and Gompertzian growth. All five can be seen as manifestations of a single microscopic model. Agent-based simulations substantiate our theory, and we can explain differences in growth curves in experimental data from em in vitro cancer cell population growth. Thus, our framework offers a possible explanation for many mean-field laws used to empirically capture seemingly unrelated cancer or microbial growth dynamics. Our results highlight that the interplay between contact inhibition and other assumptions (e.g., well-mixed) can influence our quantitative understanding of how cancer cells grow and, in turn, how they may interact.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper claims that five classical cell-population growth laws (exponential, radial, fractal, generalized logistic, and Gompertzian) are all manifestations of one microscopic model whose only interaction rule is local contact inhibition; the different macroscopic laws arise by varying auxiliary assumptions (well-mixed limit, spatial embedding, boundary conditions, etc.). Agent-based simulations are said to recover the five laws and to account for differences seen in experimental in-vitro cancer-cell growth curves.
Significance. If the mappings from the shared contact-inhibition rule to each of the five laws can be shown to be parameter-free and to follow from a single microscopic setup without law-specific auxiliary assumptions, the work would supply a biophysical unification of mean-field growth models that are currently treated as unrelated empirical fits. The agent-based validation and experimental comparison would then constitute concrete, falsifiable support for that unification.
major comments (4)
- [§3] §3 (exponential growth): the derivation invokes a well-mixed, non-spatial limit; it is not shown that this limit emerges from the same local contact-inhibition rule used for the spatially embedded cases without introducing an additional mixing parameter or rescaling.
- [§4] §4 (fractal growth): the power-law contact kernel required to obtain fractal scaling is introduced as an auxiliary assumption; the manuscript does not demonstrate that this kernel is a necessary consequence of the microscopic contact-inhibition rule rather than an independent modeling choice.
- [§5] §5 (generalized logistic and Gompertzian): the carrying-capacity term and the functional form of density-dependent inhibition appear to be chosen to match the target macroscopic equation; the text must clarify whether these forms are derived from the local rule or fitted to recover the desired law.
- [Methods] Simulation protocol (Methods): the agent-based model is reported to recover all five laws, yet the parameter values and boundary conditions used for each law are not listed; without this table it is impossible to verify that the same microscopic rule, without law-specific tuning, produces the claimed curves.
minor comments (2)
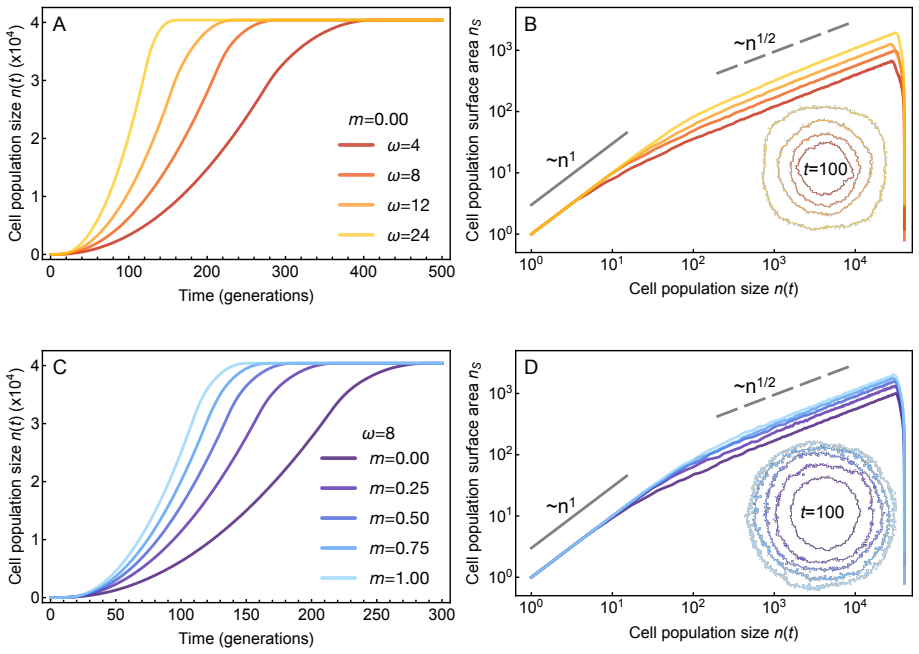
- [Figure 2] Figure 2 caption: the legend does not specify which simulation parameters correspond to each growth law.
- [§2, §4] Notation: the symbol for local contact inhibition strength is redefined between §2 and §4; a single consistent definition would improve readability.
Simulated Author's Rebuttal
We thank the referee for the careful and constructive review. The comments highlight areas where the connections between the microscopic contact-inhibition rule and the macroscopic laws, as well as the simulation details, require additional clarification. We address each point below and will revise the manuscript accordingly.
read point-by-point responses
-
Referee: [§3] §3 (exponential growth): the derivation invokes a well-mixed, non-spatial limit; it is not shown that this limit emerges from the same local contact-inhibition rule used for the spatially embedded cases without introducing an additional mixing parameter or rescaling.
Authors: We agree that the link between the well-mixed limit and the underlying local rule should be shown explicitly rather than assumed. In the revised manuscript we will add a short derivation demonstrating that the exponential law arises directly as the uniform-density limit of the same agent-based contact-inhibition dynamics when spatial correlations are neglected (i.e., when the interaction range becomes comparable to system size), without any new mixing parameter. revision: yes
-
Referee: [§4] §4 (fractal growth): the power-law contact kernel required to obtain fractal scaling is introduced as an auxiliary assumption; the manuscript does not demonstrate that this kernel is a necessary consequence of the microscopic contact-inhibition rule rather than an independent modeling choice.
Authors: The power-law kernel is presented as the effective interaction that follows when local contact inhibition is embedded in a fractal geometry. To meet the referee’s concern we will revise §4 to state explicitly that the kernel is an auxiliary modeling choice required to recover fractal scaling, and we will add a brief biophysical justification based on the local rule under fractal boundary conditions. revision: yes
-
Referee: [§5] §5 (generalized logistic and Gompertzian): the carrying-capacity term and the functional form of density-dependent inhibition appear to be chosen to match the target macroscopic equation; the text must clarify whether these forms are derived from the local rule or fitted to recover the desired law.
Authors: We will revise §5 to trace the carrying-capacity term and the specific functional form of density dependence back to the local contact-inhibition rule (via the fraction of free perimeter or available volume), making clear which steps are direct consequences of the microscopic model and which are additional assumptions needed to close the mean-field description. revision: yes
-
Referee: [Methods] Simulation protocol (Methods): the agent-based model is reported to recover all five laws, yet the parameter values and boundary conditions used for each law are not listed; without this table it is impossible to verify that the same microscopic rule, without law-specific tuning, produces the claimed curves.
Authors: We accept that a consolidated table is required for reproducibility. The revised Methods section will include a table that lists, for each growth law, the shared microscopic parameters together with the auxiliary settings (spatial embedding, boundary conditions, initial conditions) that differ across cases, thereby confirming that only the auxiliary assumptions—not the core contact-inhibition rule—are varied. revision: yes
Circularity Check
No circularity: derivations from contact-inhibition rule to macroscopic laws rely on simulations and auxiliary assumptions without reduction to fitted inputs or self-citations
full rationale
The abstract and provided excerpts present a microscopic contact-inhibition model whose outputs are substantiated by agent-based simulations rather than by algebraic identity or parameter fitting to target curves. No equations are shown that define a growth law in terms of itself or rename a fitted parameter as a prediction. Self-citations are not invoked as load-bearing uniqueness theorems. The unification claim rests on distinct auxiliary assumptions (mixing, dimensionality) applied per regime, but these are presented as external modeling choices rather than derived from the target laws. Absent explicit paper text exhibiting Eq. X = Eq. Y by construction, the derivation chain remains non-circular.
Axiom & Free-Parameter Ledger
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
λ_{i,n}=λ[1−P(ω_i|n,q_i)] … d⟨n⟩/dt=⟨λ∑[1−P(ω_i|n,q_i)]⟩−δ⟨n⟩ … radial: d⟨n⟩/dt=λd⟨n⟩^{(d−1)/d}−δ⟨n⟩; Gompertz: ˙u=−λμ_ω u ln u−δu
-
IndisputableMonolith/Foundation/AbsoluteFloorClosure.leanabsolute_floor_iff_bare_distinguishability unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
well-mixed … P(ω|n)≈(n/l)^ω … Taylor expansion … generalized logistic … Gompertz condition 2μ_ω/(σ_ω²+μ_ω²)≫|ln u|
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
A. Aguilera and B. G \'o mez-Gonz \'a lez. Genome instability: a mechanistic view of its causes and consequences. Nature Reviews Genetics, 9 0 (3): 0 204--217, 2008
work page 2008
-
[2]
P. M. Altrock, L. L. Liu, and F. Michor. The mathematics of cancer: integrating quantitative models. Nature Reviews Cancer, 15: 0 730--745, 2015
work page 2015
-
[3]
A. R. Anderson. A hybrid mathematical model of solid tumour invasion: the importance of cell adhesion. Mathematical medicine and biology: a journal of the IMA, 22 0 (2): 0 163--186, 2005
work page 2005
-
[4]
S. Benzekry, C. Lamont, A. Beheshti, A. Tracz, J. M. Ebos, L. Hlatky, and P. Hahnfeldt. Classical mathematical models for description and prediction of experimental tumor growth. PLoS Comput Biol, 10 0 (8): 0 e1003800, 2014
work page 2014
-
[5]
R. Brady and H. Enderling. Mathematical models of cancer: when to predict novel therapies, and when not to. Bulletin of mathematical biology, 81 0 (10): 0 3722--3731, 2019
work page 2019
-
[6]
R. R. Bravo, E. Baratchart, J. West, R. O. Schenck, A. K. Miller, J. Gallaher, C. D. Gatenbee, D. Basanta, M. Robertson-Tessi, and A. R. Anderson. Hybrid automata library: A flexible platform for hybrid modeling with real-time visualization. PLoS Computational Biology, 16 0 (3): 0 e1007635, 2020
work page 2020
-
[7]
I. M. Chamseddine and K. A. Rejniak. Hybrid modeling frameworks of tumor development and treatment. Wiley Interdisciplinary Reviews: Systems Biology and Medicine, 12 0 (1): 0 e1461, 2020
work page 2020
- [8]
-
[9]
H. Eagle and E. M. Levine. Growth regulatory effects of cellular interaction. Nature, 213 0 (5081): 0 1102--1106, 1967
work page 1967
-
[10]
J. Gallaher and A. R. Anderson. Evolution of intratumoral phenotypic heterogeneity: the role of trait inheritance. Interface focus, 3 0 (4): 0 20130016, 2013
work page 2013
-
[11]
P. Gerlee. The model muddle: in search of tumor growth laws. Cancer research, 73 0 (8): 0 2407--2411, 2013
work page 2013
-
[12]
C. Guiot, P. G. Degiorgis, P. P. Delsanto, P. Gabriele, and T. S. Deisboeck. Does tumor growth follow a "universal law"? Journal of Theoretical Biology, 225 0 (2): 0 147--151, 2003
work page 2003
-
[13]
H. Hatzikirou, D. Basanta, M. Simon, K. Schaller, and A. Deutsch. `go or grow': the key to the emergence of invasion in tumour progression? Mathematical medicine and biology: a journal of the IMA, 29 0 (1): 0 49--65, 2012
work page 2012
-
[14]
J. Hausser and U. Alon. Tumour heterogeneity and the evolutionary trade-offs of cancer. Nature Reviews Cancer, 20 0 (4): 0 247--257, 2020
work page 2020
- [15]
-
[16]
G. J. Kimmel, M. Dane, L. M. Heiser, P. M. Altrock, and N. Andor. Integrating mathematical modeling with high-throughput imaging explains how polyploid populations behave in nutrient-sparse environments. Cancer Research, 80 0 (22): 0 5109--5120, 2020
work page 2020
-
[17]
G. J. Kimmel, F. L. Locke, and P. M. Altrock. The roles of T cell competition and stochastic extinction events in chimeric antigen receptor t cell therapy. Proceedings of the Royal Society B, 288 0 (1947): 0 20210229, 2021
work page 1947
-
[18]
A. Laird. Dynamics of tumour growth. British Journal of Cancer, 18 0 (3): 0 490, 1964
work page 1964
-
[19]
R. Mayor and C. Carmona-Fontaine. Keeping in touch with contact inhibition of locomotion. Trends in Cell Biology, 20 0 (6): 0 319--328, 2010
work page 2010
-
[20]
A. I. McClatchey and A. S. Yap. Contact inhibition (of proliferation) redux. Current Opinion in Cell Biology, 24 0 (5): 0 685--694, 2012
work page 2012
-
[21]
J. Metzcar, Y. Wang, R. Heiland, and P. Macklin. A review of cell-based computational modeling in cancer biology. JCO Clinical Cancer Informatics, 2: 0 1--13, 2019
work page 2019
-
[22]
J. C. Mombach, N. Lemke, B. E. Bodmann, and M. A. P. Idiart. A mean-field theory of cellular growth. EPL (Europhysics Letters), 59 0 (6): 0 923, 2002
work page 2002
-
[23]
R. Orbach. Dynamics of fractal networks. Science, 231 0 (4740): 0 814--819, 1986
work page 1986
- [24]
-
[25]
V. M. P \'e rez-Garc \' a, G. F. Calvo, J. J. Bosque, O. Le \'o n-Triana, J. Jim \'e nez, J. P \'e rez-Beteta, J. Belmonte-Beitia, M. Valiente, L. Zhu, P. Garc \' a-G \'o mez, et al. Universal scaling laws rule explosive growth in human cancers. Nature Physics, pages 1--6, 2020
work page 2020
-
[26]
J. G. Reiter, M. Baretti, J. M. Gerold, A. P. Makohon-Moore, A. Daud, C. A. Iacobuzio-Donahue, N. S. Azad, K. W. Kinzler, M. A. Nowak, and B. Vogelstein. An analysis of genetic heterogeneity in untreated cancers. Nature Reviews Cancer, 19 0 (11): 0 639--650, 2019
work page 2019
-
[27]
D. Ribatti. A revisited concept: Contact inhibition of growth. from cell biology to malignancy. Experimental Cell Research, 359 0 (1): 0 17--19, 2017
work page 2017
- [28]
-
[29]
I. A. Rodriguez-Brenes, N. L. Komarova, and D. Wodarz. Tumor growth dynamics: insights into evolutionary processes. Trends in Ecology and Evolution, 28 0 (10): 0 597--604, 2013
work page 2013
-
[30]
J. A. Spratt, D. Von Fournier, J. S. Spratt, and E. E. Weber. Decelerating growth and human breast cancer. Cancer, 71 0 (6): 0 2013--2019, 1993
work page 2013
-
[31]
G. Steel. Growth kinetics of tumors. Cell population kinetics in relation to the growth and treatment of cancer, pages 59--61, 1977 a
work page 1977
-
[32]
G. G. Steel. Growth kinetics of tumors. Cell population kinetics in relation to the growth and treatment of cancer, pages 59--61, 1977 b
work page 1977
-
[33]
J. Vakkila and M. T. Lotze. Inflammation and necrosis promote tumour growth. Nature Reviews Immunology, 4 0 (8): 0 641--648, 2004
work page 2004
-
[34]
S. T. Vittadello, S. W. McCue, G. Gunasingh, N. K. Haass, and M. J. Simpson. Examining go-or-grow using fluorescent cell-cycle indicators and cell-cycle-inhibiting drugs. Biophysical Journal, 2020
work page 2020
-
[35]
J. West and P. Newton. Cellular interactions constrain tumor growth. Proceedings of the National Academy of Sciences, 116 0 (6): 0 1918--1923, 2019. ISSN 0027-8424. doi:10.1073/pnas.1804150116
-
[36]
J. West, Z. Hasnain, P. Macklin, and P. K. Newton. An evolutionary model of tumor cell kinetics and the emergence of molecular heterogeneity driving gompertzian growth. SIAM Review, 58 0 (4): 0 716--736, 2016
work page 2016
-
[37]
T. E. Wheldon. Mathematical models in cancer research. Taylor & Francis, 1988
work page 1988
-
[38]
D. Wodarz and N. Komarova. Computational biology of cancer: Lecture notes and mathematical modeling. World Scientific Publishing, 2005
work page 2005
-
[39]
M. Zwietering, I. Jongenburger, F. Rombouts, and K. Van't Riet. Modeling of the bacterial growth curve. Applied and Environmental Microbiology, 56 0 (6): 0 1875--1881, 1990
work page 1990
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.