A lattice structure for ancestral configurations arising from the relationship between gene trees and species trees
Pith reviewed 2026-05-24 13:07 UTC · model grok-4.3
The pith
For matching gene and species tree topologies, paths on the ancestral configuration digraph stand in bijection with labeled histories.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
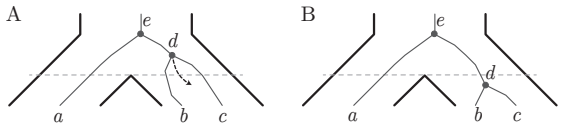
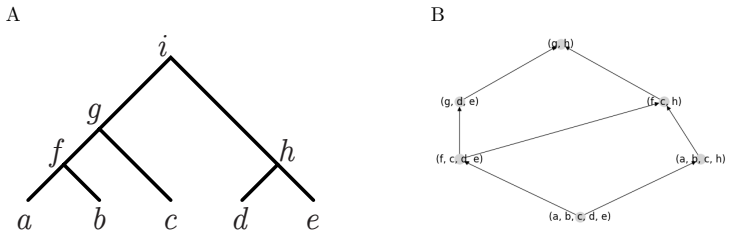
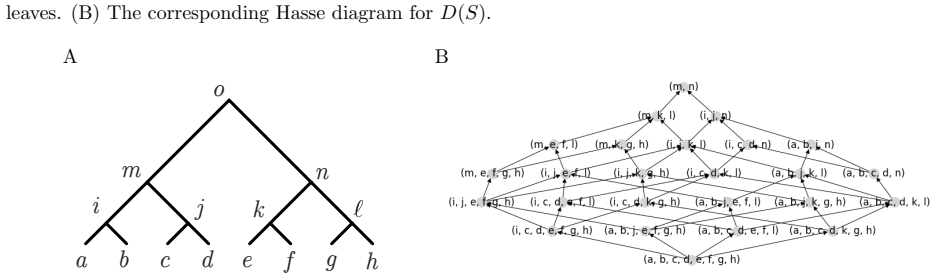
When gene tree topology G equals species tree topology S, a specific set of paths on the digraph of ancestral configurations is in bijection with the set of labeled histories.
What carries the argument
The digraph of ancestral configurations, obtained from the tree topology by iterated Cartesian products of graphs, whose selected paths enumerate labeled histories.
If this is right
- Closed-form expressions become available for the number of labeled histories in several infinite families of trees.
- Pairs of topologies (G,S) that attain the maximum number of ancestral configurations for fixed G can be characterized.
- Enumeration of other combinatorial features of gene-species tree pairs becomes possible through path counting on the same digraphs.
Where Pith is reading between the lines
- The path-counting method may yield polynomial-time algorithms for labeled-history enumeration on trees of moderate size.
- Analogous lattice constructions could organize other ordered coalescence objects such as ranked tree shapes.
- The graphical representation may support visualization tools that display coalescence orderings directly on the lattice.
Load-bearing premise
The iterated Cartesian product construction produces a digraph whose vertices and edges correctly encode the ancestral configurations whenever the gene-tree and species-tree topologies are identical.
What would settle it
For any small matching tree pair whose number of labeled histories is already known by direct enumeration, a mismatch between that number and the number of qualifying paths in the constructed digraph.
Figures












read the original abstract
To a given gene tree topology $G$ and species tree topology $S$ with leaves labeled bijectively from a fixed set $X$, one can associate a set of ancestral configurations, each of which encodes a set of gene lineages that can be found at a given node of the species tree. We introduce a lattice structure on ancestral configurations, studying the directed graphs that provide graphical representations of lattices of ancestral configurations. For a matching gene tree topology and species tree topology $G=S$, we present a method for defining the digraph of ancestral configurations from the tree topology by using iterated cartesian products of graphs. We show that a specific set of paths on the digraph of ancestral configurations is in bijection with the set of labeled histories -- a well-known phylogenetic object that enumerates possible temporal orderings of the coalescences of a tree. For each of a series of tree families, we obtain closed-form expressions for the number of labeled histories by using this bijection to count paths on associated digraphs. Finally, we prove that our lattice construction extends to nonmatching tree pairs, and we use it to characterize pairs $(G,S)$ having the maximal number of ancestral configurations for a fixed $G$. We discuss how the construction provides new methods for performing enumerations of combinatorial aspects of gene and species trees.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript associates ancestral configurations to a gene tree topology G and species tree topology S on the same leaf set. It equips these configurations with a lattice structure whose Hasse diagram is realized as a digraph. For the special case G = S the authors construct the digraph by iterated Cartesian products of smaller graphs derived from the tree topology, prove that a distinguished subset of paths in this digraph is in bijection with the labeled histories of the tree, and obtain closed-form expressions for the number of labeled histories on several infinite families of trees. The lattice construction is then extended to G ≠ S and used to characterize pairs that maximize the number of ancestral configurations for fixed G.
Significance. If the bijection and the Cartesian-product construction are valid, the work supplies a new, graph-theoretic route to enumerating labeled histories—an object central to coalescent theory and phylogenetic combinatorics. The closed forms for concrete tree families and the characterization of maximal (G,S) pairs are concrete, falsifiable contributions. The lattice formalism itself may also furnish new tools for comparing gene and species trees.
major comments (1)
- [method for defining the digraph (paragraph following the lattice definition)] The central claim that the iterated Cartesian product yields a digraph whose distinguished paths are in bijection with labeled histories rests on the construction correctly encoding the partial order of coalescences. For unbalanced topologies the product treats sub-configurations as independent factors, yet global coalescence constraints may produce extraneous vertices or omit valid lineage sets. A concrete verification for at least one unbalanced matching pair (e.g., the caterpillar with four or five leaves) is needed to confirm that the bijection survives; the abstract and method description do not supply this check.
minor comments (2)
- Notation for the Cartesian product of graphs and for the distinguished path set should be introduced with an explicit small example (e.g., the balanced quartet) before the general statement.
- The closed-form expressions are stated for several families; a short table listing the families, the formulas, and the first few numerical values would improve readability.
Simulated Author's Rebuttal
We thank the referee for their careful reading of the manuscript and for identifying the need for an explicit check on unbalanced topologies. We address the single major comment below.
read point-by-point responses
-
Referee: [method for defining the digraph (paragraph following the lattice definition)] The central claim that the iterated Cartesian product yields a digraph whose distinguished paths are in bijection with labeled histories rests on the construction correctly encoding the partial order of coalescences. For unbalanced topologies the product treats sub-configurations as independent factors, yet global coalescence constraints may produce extraneous vertices or omit valid lineage sets. A concrete verification for at least one unbalanced matching pair (e.g., the caterpillar with four or five leaves) is needed to confirm that the bijection survives; the abstract and method description do not supply this check.
Authors: The recursive definition of the iterated Cartesian product follows the tree hierarchy directly: at each internal node the product is formed from the digraphs of its child subtrees, with edges between factors only when the corresponding coalescences are compatible under the species-tree partial order. This encoding ensures global constraints are respected even when subtrees are unbalanced, because the product order is induced by the ancestor-descendant relations in the full topology rather than by treating factors as fully independent. Nevertheless, we agree that an explicit small-case verification would strengthen the presentation. In the revised manuscript we will insert, immediately after the general construction, a fully worked example for the 5-leaf caterpillar (an unbalanced matching pair). The example will display the digraph, list all distinguished paths, and confirm that their number equals the known count of labeled histories (42) with no extraneous vertices or omitted lineage sets. revision: yes
Circularity Check
No circularity: combinatorial bijection and path-counting construction is self-contained
full rationale
The paper defines ancestral configurations via iterated Cartesian products of graphs from the tree topology, then proves a bijection between specific paths on the resulting digraph and labeled histories. Closed-form counts for tree families follow directly from this bijection. No parameters are fitted, no self-citations are load-bearing for the central claim, and no step reduces a derived quantity to its own input by definition. The construction is a standard combinatorial enumeration independent of the target counts.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption Ancestral configurations form a partially ordered set that is a lattice under the natural inclusion or coalescence order.
- standard math Cartesian product of graphs preserves the required path-counting properties for building the digraph from tree topology.
Reference graph
Works this paper leans on
-
[1]
Aho, A. V. and Sloane, N. J. A. 1973. Some doubly exponential sequences, Fibonacci Quarterly 11, 429–437
work page 1973
-
[2]
Alimpiev, E. and Rosenberg, N. A. 2021. Enumeration of coalescent histories for caterpillar species trees and p-pseudocaterpillar gene trees, Advances in Applied Mathematics 131, 102265
work page 2021
-
[3]
Bienvenu, F., Lambert, A., and Steel, M. 2022. Combinatorial and stochastic properties of ranked tree-child networks, Random Structures & Algorithms 60 (4), 653–689
work page 2022
-
[4]
Brown, J. K. 1994. Probabilities of evolutionary trees, Systematic Biology 43 (1), 78–91
work page 1994
-
[5]
Colijn, C. and Plazzotta, G. 2018. A metric on phylogenetic tree shapes, Systematic Biology 67, 113–126
work page 2018
-
[6]
Degnan, J. H. and Rosenberg, N. A. 2009. Gene tree discordance, phylogenetic inference and the multispecies coalescent, Trends in Ecology and Evolution 24, 332–340
work page 2009
-
[7]
Degnan, J. H., Rosenberg, N. A., and Stadler, T. 2012. The probability distribution of ranked gene trees on a species tree, Mathematical Biosciences 235 (1), 45–55
work page 2012
-
[8]
Degnan, J. H. and Salter, L. A. 2005. Gene tree distributions under the coalescent process, Evolution 59 (1), 24–37
work page 2005
-
[9]
Disanto, F., Fuchs, M., Huang, C.-Y., Paningbatan, A. R., and Rosenberg, N. A. 2024. The distributions under two species-tree models of the total number of ancestral configurations for matching gene trees and species trees, Advances in Applied Mathematics 152, 102594
work page 2024
-
[10]
Disanto, F., Fuchs, M., Paningbatan, A. R., and Rosenberg, N. A. 2022. The distributions under two species-tree models of the number of root ancestral configurations for matching gene trees and species trees, Annals of Applied Probability 32 (6), 4426–4458
work page 2022
-
[11]
Disanto, F. and Rosenberg, N. A. 2015. Coalescent histories for lodgepole species trees, Journal of Computational Biology 22 (10), 918–929
work page 2015
-
[12]
Disanto, F. and Rosenberg, N. A. 2016. Asymptotic properties of the number of matching coalescent histories for caterpillar-like families of species trees, IEEE/ACM Transactions on Computational Biology and Bioinformatics 13 (5), 913–925
work page 2016
-
[13]
Disanto, F. and Rosenberg, N. A. 2017. Enumeration of ancestral configurations for matching gene trees and species trees, Journal of Computational Biology 24 (9), 831–850
work page 2017
-
[14]
Introduction to Lattice Theory with Computer Science Applications
Garg, V. K. 2015. “Introduction to Lattice Theory with Computer Science Applications”, John Wiley & Sons, New York. Gr¨ atzer, G. 2011. “Lattice Theory: Foundation”, Springer, Basel
work page 2015
-
[15]
Harding, E. 1971. The probabilities of rooted tree-shapes generated by random bifurcation, Advances in Applied Probability 3 (1), 44–77
work page 1971
-
[16]
Himwich, Z. M. and Rosenberg, N. A. 2020. Roadblocked monotonic paths and the enumeration of coalescent histories for non-matching caterpillar gene trees and species trees, Advances in Applied Mathematics 113, 101939
work page 2020
-
[17]
King, M. C. and Rosenberg, N. A. 2023. A mathematical connection between single-elimination sports tournaments and evolutionary trees, Mathematics Magazine in press
work page 2023
-
[18]
Mathur, S. and Rosenberg, N. A. 2023. All galls are divided into three or more parts: recursive enumeration of labeled histories for galled trees, Algorithms for Molecular Biology 18, 1
work page 2023
-
[19]
A., Bhaskar, A., Disanto, F., and Rosenberg, N
Palacios, J. A., Bhaskar, A., Disanto, F., and Rosenberg, N. A. 2022. Enumeration of binary trees compatible with a perfect phylogeny, Journal of Mathematical Biology 84 (6), 1–37. 20
work page 2022
-
[20]
Rosenberg, N. A. 2007. Counting coalescent histories, Journal of Computational Biology 14, 360–377
work page 2007
-
[21]
Rosenberg, N. A. 2013. Coalescent histories for caterpillar-like families, IEEE/ACM Transactions on Computational Biology and Bioinformatics 10 (5), 1253–1262
work page 2013
-
[22]
Rosenberg, N. A. 2019. Enumeration of lonely pairs of gene trees and species trees by means of antipodal cherries, Advances in Applied Mathematics 102, 1–17
work page 2019
-
[23]
Rosenberg, N. A. 2021. On the Colijn–Plazzotta numbering scheme for unlabeled binary rooted trees, Discrete Applied Mathematics 291, 88–98
work page 2021
-
[24]
Sabidussi, G. 1959. Graph multiplication, Mathematische Zeitschrift 72 (1), 446–457
work page 1959
-
[25]
Song, Y. S., Lyngso, R., and Hein, J. 2006. Counting all possible ancestral configurations of sample sequences in population genetics, IEEE/ACM Transactions on Computational Biology and Bioinformatics 3 (3), 239–251
work page 2006
-
[26]
Stadler, T. and Degnan, J. H. 2012. A polynomial time algorithm for calculating the probability of a ranked gene tree given a species tree, Algorithms for Molecular Biology 7 (1), 7
work page 2012
-
[27]
Enumerative Combinatorics, Volume 1
Stanley, R. P. 2011. “Enumerative Combinatorics, Volume 1”, Cambridge University Press, Cambridge, UK, second edition
work page 2011
-
[28]
Phylogeny: Discrete and Random Processes in Evolution
Steel, M. 2016. “Phylogeny: Discrete and Random Processes in Evolution”, SIAM, Philadelphia
work page 2016
-
[29]
Steel, M. and McKenzie, A. 2001. Properties of phylogenetic trees generated by Yule-type speciation models, Mathematical Biosciences 170 (1), 91–112
work page 2001
-
[30]
Probabilistic Structures in Evolution
Wiehe, T. 2021. Counting, grafting and evolving binary trees. in “Probabilistic Structures in Evolution.” (E. Baake and A. Wakolbinger, eds), p. 427–450, Zurich, EMS Publishing House
work page 2021
-
[31]
Wu, Y. 2012. Coalescent-based species tree inference from gene tree topologies under incomplete lineage sorting by maximum likelihood, Evolution 66 (3), 763–775
work page 2012
-
[32]
Wu, Y. 2016. An algorithm for computing the gene tree probability under the multispecies coalescent and its application in the inference of population tree, Bioinformatics 32 (12), i225–i233. 21
work page 2016
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.