Evolutionary origin of the bipartite architecture of dissipative cellular networks
Pith reviewed 2026-05-23 19:26 UTC · model grok-4.3
The pith
Dissipative biological networks evolve a decoupled fuel module even when selected only for function.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
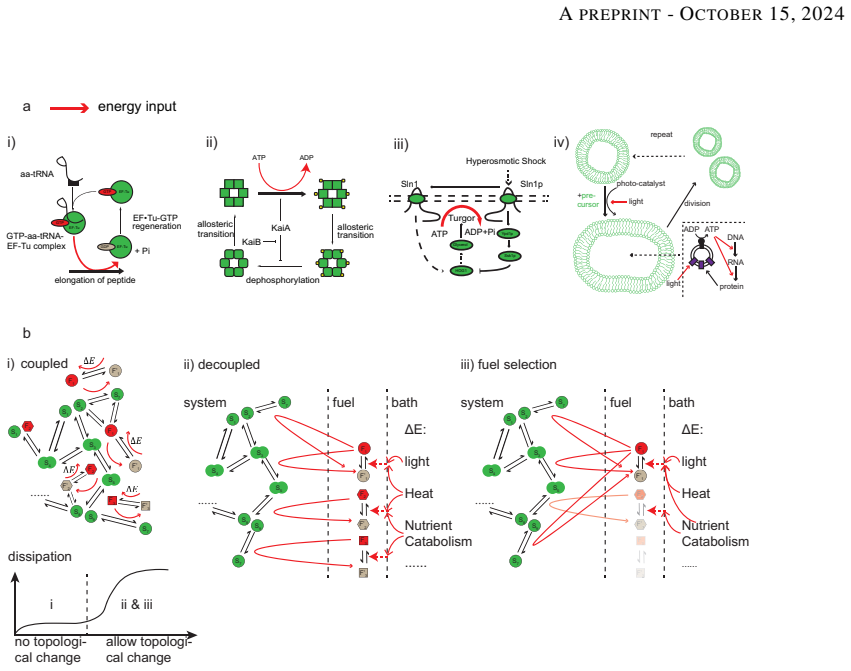
Through evolutionary simulations on premature networks of kinetic proofreading, activator-inhibitor oscillators, and two adaptive-response models, with selection imposed solely on network function, the networks evolve to decouple high-energy molecules as fuels from the functional module. This fuel decoupling produces higher overall dissipation, increases robustness to parameter or structure perturbations, and in most cases improves performance. Theoretical analysis of kinetic proofreading and general energy-driven networks confirms that decoupling guarantees higher dissipation while supporting better function.
What carries the argument
Fuel decoupling, the evolutionary isolation of high-energy-molecule reactions into a separate module from the functional reactions in dissipative networks.
If this is right
- Higher overall energy dissipation is achieved compared with coupled architectures.
- Robustness to parameter and structural perturbations increases.
- Performance improves in the majority of dissipative network cases examined.
- The bipartite structure arises as a side effect of selection on function alone.
Where Pith is reading between the lines
- Synthetic biology designs could deliberately insert fuel decoupling to raise efficiency and stability.
- The pattern may appear across other classes of energy-dissipating networks not tested here.
- Decoupling could reduce the number of evolutionary trials needed to reach functional networks.
- Cellular pathways that already show separated energy inputs may reflect this same evolutionary pressure.
Load-bearing premise
The evolutionary simulations select solely on network function and the observed decoupling is not produced by the specific kinetic models or parameter choices used.
What would settle it
An evolutionary simulation in which a dissipative network reaches high functional performance without separating fuel reactions, or a real cellular network that maintains high function while keeping fuel and functional reactions tightly coupled.
Figures





read the original abstract
Recently, plenty research has been done on discovering the role of energy dissipation in biological networks, most of which focus on the relationship of dissipation and functionality. However, the development of networks science urged us to fathom the systematic architecture of biological networks and their evolutionary advantages. We found the dissipation of biological dissipative networks is highly related to their structure. By interrogating these well-adapted networks, we find that the energy producing module is relatively isolated in all situations. We applied evolutionary simulation and analysis on premature networks of classic dissipative networks, namely kinetic proofreading, activator-inhibitor oscillator and two typical adaptative response models. We found despite that selection was imposed merely on the network function, the networks tended to decouple high energy molecules as fuels from the functional module, to achieve higher overall dissipation during the course of evolution. Furthermore, we find that decoupled fuel modules can increase the robustness of the networks towards parameter or structure perturbations. We provide theoretical analysis on the kinetic proofreading networks and the general case of energy-driven networks. We find fuel decoupling can guarantee higher dissipation and, in most cases when considering dissipative networks, higher performance. We conclude that fuel decoupling is an evolutionary outcome and bears benefits during evolution.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript claims that evolutionary simulations on premature networks of kinetic proofreading, activator-inhibitor oscillators, and two adaptive-response models show that selection imposed only on functional performance (error rate, oscillation amplitude, adaptation accuracy) drives decoupling of high-energy fuel modules from the functional core. This structural change is reported to increase overall dissipation, robustness to parameter and structural perturbations, and often performance. Theoretical analysis on the kinetic proofreading case and the general energy-driven network case is presented to argue that fuel decoupling guarantees higher dissipation.
Significance. If the central claim holds after verification of generality, the work would establish a mechanistic link between dissipative network architecture and evolutionary selection on function, explaining the prevalence of bipartite structures in biological networks and identifying robustness benefits. The use of multiple distinct models plus theory is a positive feature, though the result's independence from specific kinetic assumptions remains to be demonstrated.
major comments (3)
- [theoretical analysis on kinetic proofreading networks] Theoretical analysis section on kinetic proofreading: the assertion that decoupling 'guarantees higher dissipation' must be shown to be independent of the Michaelis-Menten rate forms employed in the simulations; if the derivation relies on the same kinetic assumptions, the guarantee reduces to a property of the model rather than a general consequence of decoupling.
- [evolutionary simulation and analysis on premature networks] Evolutionary simulation results for activator-inhibitor oscillator: the reported evolution toward fuel decoupling under selection on oscillation amplitude should be tested against alternative rate laws (e.g., mass-action kinetics instead of the Hill functions used); without this control, the structural outcome may be an artifact of the chosen functional forms rather than a general outcome of selection on function.
- [theoretical analysis on the general case of energy-driven networks] General case of energy-driven networks: the claim that decoupling yields higher performance 'in most cases' requires an explicit statement of the conditions and the set of dissipative networks considered; the two adaptive-response ODE systems alone do not establish the scope of the 'most cases' statement.
minor comments (2)
- The abstract and methods should include a precise definition of 'fuel decoupling' with reference to the network diagrams or adjacency matrices used in each model.
- Reproducibility requires reporting of evolutionary algorithm parameters (population size, mutation rate, selection strength, number of independent runs) for all four models.
Simulated Author's Rebuttal
We thank the referee for the constructive comments, which help clarify the scope and generality of our results. We address each major comment below and indicate planned revisions.
read point-by-point responses
-
Referee: Theoretical analysis section on kinetic proofreading: the assertion that decoupling 'guarantees higher dissipation' must be shown to be independent of the Michaelis-Menten rate forms employed in the simulations; if the derivation relies on the same kinetic assumptions, the guarantee reduces to a property of the model rather than a general consequence of decoupling.
Authors: The derivation for kinetic proofreading establishes higher dissipation from the structural separation of the fuel module, relying on the general properties of energy dissipation in enzymatic networks (free energy drop across the decoupled step) rather than specific parameter values or functional forms of the rate laws. The proof uses the inequality between total dissipation and the functional module's contribution, which holds as long as the fuel production is isolated. To strengthen clarity, we will add an explicit paragraph stating the minimal assumptions (irreversible fuel production and no direct back-coupling) and note that the result is independent of Michaelis-Menten details. revision: yes
-
Referee: Evolutionary simulation results for activator-inhibitor oscillator: the reported evolution toward fuel decoupling under selection on oscillation amplitude should be tested against alternative rate laws (e.g., mass-action kinetics instead of the Hill functions used); without this control, the structural outcome may be an artifact of the chosen functional forms rather than a general outcome of selection on function.
Authors: We acknowledge that the original simulations used Hill functions for the oscillator. We have now run control evolutionary simulations replacing Hill functions with mass-action kinetics for the core reactions while retaining the same selection criterion on amplitude. Fuel decoupling still emerges as the dominant evolutionary outcome, with comparable increases in dissipation and robustness. These additional results will be included in a revised supplementary figure and discussed in the main text. revision: yes
-
Referee: General case of energy-driven networks: the claim that decoupling yields higher performance 'in most cases' requires an explicit statement of the conditions and the set of dissipative networks considered; the two adaptive-response ODE systems alone do not establish the scope of the 'most cases' statement.
Authors: The 'most cases' claim originates from the general theoretical analysis of energy-driven networks (Section on general case), which considers any dissipative network possessing a separable high-energy fuel module whose production can be decoupled without altering the functional core. The two adaptive-response models serve as concrete illustrations, not as the full set. We will revise the text to explicitly delineate the conditions (networks with at least one energy-consuming step that can be isolated, positive dissipation requirement, and functional selection on output accuracy) and state that the result applies to the broad class of such networks rather than being limited to the examples shown. revision: yes
Circularity Check
No circularity; theoretical guarantee and simulations remain independent of fitted inputs
full rationale
The abstract and skeptic summary describe evolutionary simulations that select solely on functional metrics (error rate, oscillation amplitude, adaptation accuracy) across standard models (Michaelis-Menten kinetic proofreading, Hill-function oscillators, adaptive ODEs), followed by a separate theoretical analysis concluding that fuel decoupling guarantees higher dissipation. No quoted equations, self-citations, or fitted parameters are supplied that would reduce the dissipation guarantee to a definition or to the simulation fitness function itself. The derivation chain therefore does not exhibit any of the enumerated circular patterns; the result is not forced by construction from the inputs.
Axiom & Free-Parameter Ledger
Reference graph
Works this paper leans on
-
[1]
Evan Eisenberg and Terrell L. Hill. Muscle contraction a nd free energy transduction in biological systems. Science, 227(4690):999–1006, 1985
work page 1985
-
[2]
Terrell L. Hill. Free Energy Transduction and Biochemical Cycle Kinetics . Springer New Y ork : Imprint: Springer, New Y ork, NY , 1st 1989. edition, 1989
work page 1989
-
[3]
Daniel A. Beard and Hong Qian. Chemical biophysics : quantitative analysis of cellular sy stems. Cambridge texts in biomedical engineering. Cambridge University Pre ss, Cambridge ; New Y ork, 2008
work page 2008
-
[4]
J. J. Hopfield. Kinetic proofreading: A new mechanism for reducing errors in biosynthetic processes requiring high specificity. Proceedings of the National Academy of Sciences , 71(10):4135–4139, 1974
work page 1974
-
[5]
Y . S. Cao, H. L. Wang, Q. Ouyang, and Y . H. Tu. The free-ener gy cost of accurate biochemical oscillations. Nature Physics, 11(9):772–+, 2015. Cq6iy Times Cited:89 Cited References Count:43
work page 2015
-
[6]
G. Lan, P . Sartori, S. Neumann, V . Sourjik, and Y . H. Tu. Th e energy-speed-accuracy trade-off in sensory adaptation. Nature Physics, 8(5):422–428, 2012. 936of Times Cited:248 Cited Referenc es Count:43. 11 A PREPRINT - O CTOBER 15, 2024
work page 2012
-
[7]
Wenzhe Ma, Ala Trusina, Hana El-Samad, Wendell A. Lim, an d Chao Tang. Defining network topologies that can achieve biochemical adaptation. Cell, 138(4):760–773, 2009
work page 2009
-
[8]
Robert Marsland and Jeremy L. England. Active regenerat ion unites high- and low-temperature features in cooperative self-assembly. Physical Review E, 98(2):022411, 2018. PRE
work page 2018
-
[9]
S. Rasmussen, A. Constantinescu, and C. Svaneborg. Gene rating minimal living systems from non-living ma- terials and increasing their evolutionary abilities. Philosophical Transactions of the Royal Society B-Biologi cal Sciences, 371(1701), 2016. Dx5zf Times Cited:18 Cited References Co unt:64
work page 2016
-
[10]
P . Adamski, M. Eleveld, A. Sood, A. Kun, A. Szilagyi, T. C zaran, E. Szathmary, and S. Otto. From self- replication to replicator systems en route to de novo life. Nature Reviews Chemistry, 4(8):386–403, 2020. Mv3eq Times Cited:37 Cited References Count:148
work page 2020
-
[11]
C. Xu, S. Hu, and X. Y . Chen. Artificial cells: from basic s cience to applications. Materials T oday, 19(9):516– 532, 2016. Ef8xa Times Cited:171 Cited References Count:15 2
work page 2016
-
[12]
DeClue, Pierre-Alain Monnard, James A
Michael S. DeClue, Pierre-Alain Monnard, James A. Bail ey, Sarah E. Maurer, Gavin E. Collis, Hans-Joachim Ziock, Steen Rasmussen, and James M. Boncella. Nucleobase m ediated, photocatalytic vesicle formation from an ester precursor. Journal of the American Chemical Society , 131(3):931–933, 2009. doi: 10.1021/ja808200n
-
[13]
W . Y u, K. Sato, M. Wakabayashi, T. Nakaishi, E. P . Ko-Mitamura, Y . Shima, I. Urabe, and T. Y omo. Synthesis of functional protein in liposome. Journal of Bioscience and Bioengineering , 92(6):590–593, 2001. 522ed Times Cited:173 Cited References Count:21
work page 2001
-
[14]
S. Berhanu, T. Ueda, and Y . Kuruma. Artificial photosynthetic cell producing energy for protein synthesis. Nature Communications, 10, 2019. Hp9ei Times Cited:171 Cited References Count:37
work page 2019
-
[15]
A. Wolos, R. Roszak, A. Zadlo-Dobrowolska, W . Beker, B. Mikulak-Klucznik, G. Spolnik, M. Dygas, S. Szymkuc, and B. A. Grzybowski. Synthetic connectivity, e mergence, and self-regeneration in the network of prebiotic chemistry. Science, 369(6511):1584–+, 2020. Nv9td Times Cited:47 Cited Refer ences Count:162
work page 2020
-
[16]
Jordan M. Horowitz and Jeremy L. England. Spontaneous fi ne-tuning to environment in many-species chemical reaction networks. Proceedings of the National Academy of Sciences , 114(29):7565–7570, 2017
work page 2017
-
[17]
P . Sassone-Corsi. The cyclic amp pathway. Cold Spring Harb Perspect Biol , 4(12), 2012. 1943-0264 Sassone- Corsi, Paolo Journal Article Review United States 2012/12/ 05 Cold Spring Harb Perspect Biol. 2012 Dec 1;4(12):a011148. doi: 10.1101/cshperspect.a011148
-
[18]
S. E. Irving, N. R. Choudhury, and R. M. Corrigan. The str ingent response and physiological roles of (pp)pgpp in bacteria. Nat Rev Microbiol , 19(4):256–271, 2021. 1740-1534 Irving, Sophie E Orcid: 00 00-0003-0772- 1946 Choudhury, Naznin R Orcid: 0000-0002-7266-5736 Corri gan, Rebecca M Orcid: 0000-0002-6031-1148 104110/Z/14/Z/WT/Wellcome Trust/United Ki...
-
[19]
H. Qian. Reducing intrinsic biochemical noise in cells and its thermodynamic limit. Journal of Molecular Biology, 362(3):387–392, 2006. 089md Times Cited:38 Cited Referen ces Count:27
work page 2006
-
[20]
T. W . Traut. Physiological concentrations of purines a nd pyrimidines. Molecular and Cellular Biochemistry , 140(1):1–22, 1994. Pv684 Times Cited:1272 Cited Reference s Count:107
work page 1994
-
[21]
Agnes Szejka and Barbara Drossel. Evolution of boolean networks under selection for a robust response to external inputs yields an extensive neutral space. Physical Review E, 81(2):021908, 2010. PRE
work page 2010
-
[22]
J. V . Greiner and T. Glonek. Intracellular atp concentr ation and implication for cellular evolution. Biology-Basel, 10(11), 2021. Xe5bm Times Cited:7 Cited References Count:9 6
work page 2021
-
[23]
Avinash Patel, Liliana Malinovska, Shambaditya Saha, Jie Wang, Simon Alberti, Y amuna Krishnan, and An- thony A. Hyman. Atp as a biological hydrotrope. Science, 356(6339):753–756, 2017. 12
work page 2017
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.