Bistability of travelling waves and wave-pinning states in a mass-conserved reaction-diffusion system: From bifurcations to implications for actin waves
Pith reviewed 2026-05-23 19:22 UTC · model grok-4.3
The pith
In mass-conserved models of Rho-GTPase and F-actin, moderate domain lengths create bistability between wave-pinning states and travelling waves.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
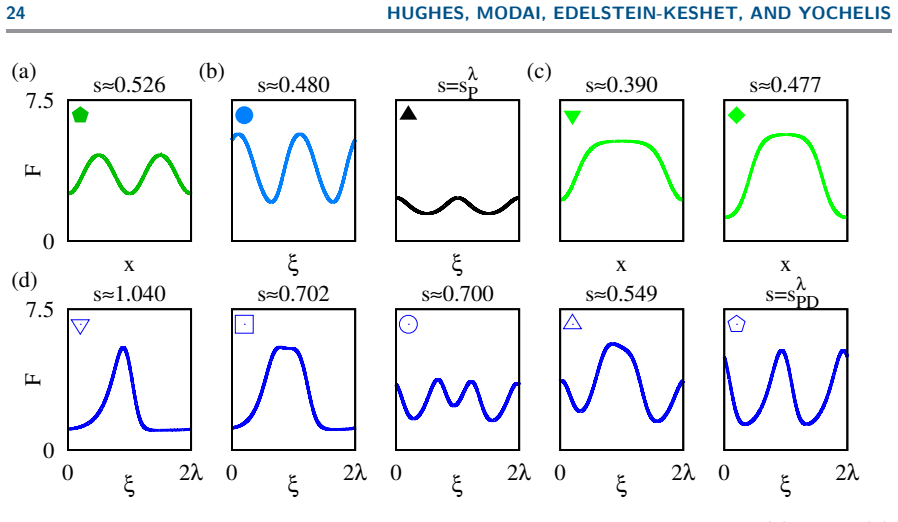
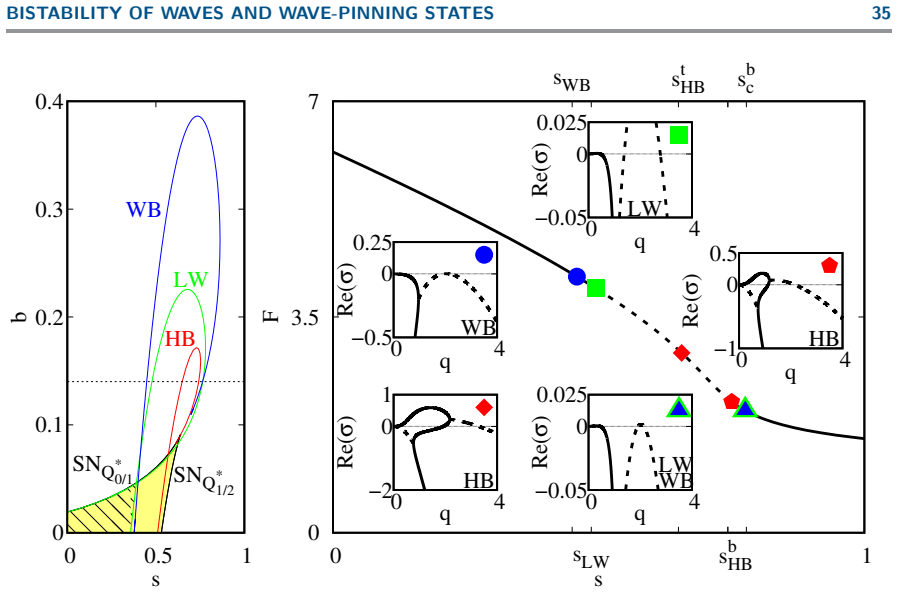
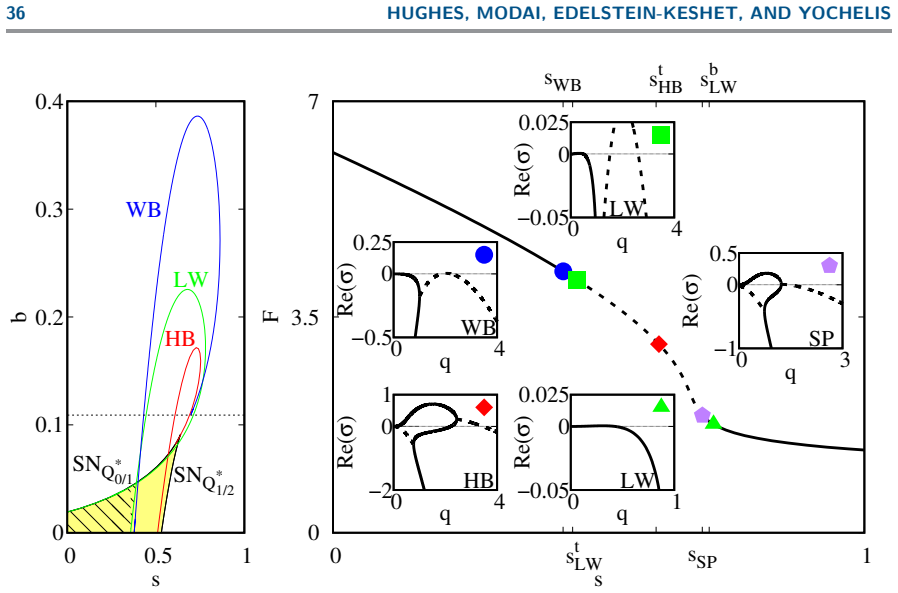
The codimension-2 Hopf instability organizes a family of solutions distinguished by mass-conservation regimes and classified by domain size: steady wave-pinning states (mesas obeying the Maxwell construction), propagating fronts and excitable pulses, and travelling and standing waves; in particular, bistability between wave-pinning and travelling waves unfolds through domain length on moderate domains.
What carries the argument
Codimension-2 long-wavelength and finite-wavenumber Hopf instability on the mass-conserved 1D periodic domain.
Load-bearing premise
The specific negative-feedback loop in which F-actin inactivates Rho-GTPase while Rho-GTPase promotes F-actin, together with strict mass conservation, holds on the 1D periodic domain.
What would settle it
Numerical continuation or simulation on moderate domains showing that the region of bistability between wave-pinning and travelling waves does not unfold with domain length.
Figures






















read the original abstract
Eukaryotic cells demonstrate a wide variety of dynamic patterns of filamentous actin (F-actin) and its regulators. Some of these patterns play important roles in cell functions, such as distinct motility modes, which motivate this study. We devise a mass-conserved reaction-diffusion model for active and inactive Rho-GTPase and F-actin in the cell cortex. The mass-conserved Rho-GTPase system promotes F-actin, which feeds back to inactivate the former. We study the model on a 1D periodic domain (edge of thin sheet-like cell) using bifurcation theory in the framework of spatial dynamics, complemented with numerical simulations. Among several discussed bifurcations, the analysis centers on the study of the codimension-2 long wavelength and finite wavenumber Hopf instability, in which we describe a rich structure of steady wave-pinning states (a.k.a. mesas, obeying the Maxwell construction), propagating coherent solutions (fronts and excitable pulses), and travelling and standing waves, all distinguished by mass conservation regimes and classified by domain sizes. Specifically, we highlight the unexpected conditions for bistability between steady wave-pinning and travelling wave states on moderate domain sizes, i.e., unfolding through domain length. These results uncover and exemplify possible mechanisms of coexistence, robustness, and transitions between distinct cellular motility modes, including directed migration, turning, and ruffling. More broadly, the results indicate that non-gradient reaction-diffusion models comprising mass conservation have distinct pattern formation mechanisms that motivate further investigations, such as the unfolding of codimension-3 instabilities and T-points.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript presents a mass-conserved reaction-diffusion model coupling active/inactive Rho-GTPase with F-actin, where the Rho system promotes F-actin and F-actin inactivates Rho-GTPase. On a 1D periodic domain, spatial-dynamics bifurcation analysis (centered on the codim-2 long-wavelength/finite-wavenumber Hopf point) combined with numerical simulations classifies steady wave-pinning (mesa) states obeying the Maxwell construction, fronts, excitable pulses, travelling/standing waves, and identifies bistability between wave-pinning and travelling waves that unfolds with domain length at moderate sizes, all distinguished by mass-conservation regimes.
Significance. If the central bifurcation results hold, the work demonstrates how mass conservation in non-gradient RD systems produces distinct mechanisms for pattern coexistence and transitions, with direct implications for cellular motility modes (directed migration, turning, ruffling). The explicit classification of solutions by domain size via spatial dynamics, together with the identification of the codim-2 unfolding, constitutes a clear advance over generic Turing or wave-pinning analyses.
minor comments (3)
- [Abstract] Abstract: the description of the feedback loop and the codim-2 point would be strengthened by a single displayed model equation or parameter list so that the Maxwell-construction claim can be immediately verified against the conservation law.
- [Numerical methods] The numerical continuation results that confirm the bistability region on moderate domains should include a brief statement of the discretization scheme, tolerance, and how the periodic boundary conditions are enforced, to allow reproduction of the domain-length unfolding.
- [Figures] Figure captions for the bifurcation diagrams should explicitly label which branches correspond to the wave-pinning versus travelling-wave states and indicate the mass-conservation regime for each panel.
Simulated Author's Rebuttal
We thank the referee for their positive summary and significance assessment of our work. The recommendation for minor revision is noted; we will prepare a revised manuscript accordingly. No specific major comments were raised in the report.
Circularity Check
No significant circularity
full rationale
The derivation applies standard spatial-dynamics bifurcation analysis (codim-2 long-wavelength/finite-wavenumber Hopf point) and direct numerical simulation to the stated mass-conserved RD system on a 1D periodic domain. All reported states (wave-pinning mesas obeying Maxwell construction, fronts, pulses, travelling/standing waves) are obtained from the model's PDEs and boundary conditions without any fitted parameter being relabeled as a prediction, without self-definitional closure, and without load-bearing reliance on prior self-citations whose validity is presupposed. The bistability result is an output of the unfolding with domain length, not an input. The analysis is therefore self-contained against external benchmarks.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption The Rho-GTPase system is strictly mass-conserved and promotes F-actin which in turn inactivates Rho-GTPase.
- domain assumption Analysis is performed on a 1D periodic domain representing the edge of a thin sheet-like cell.
Reference graph
Works this paper leans on
-
[1]
J. Algorta, A. Fele-Paranj, J. M. Hughes, and L. Edelstein-Keshet , Modeling and simulating single and collective cell motility , Cold Spring Harb. Perspect. in Biol., (2025)
work page 2025
-
[2]
J. Allard and A. Mogilner , Traveling waves in actin dynamics and cell motility , Curr. Opin. Cell Biol., 25 (2013), pp. 107–115
work page 2013
-
[3]
Y. Arai, T. Shibata, S. Matsuoka, M. J. Sato, T. Yanagida, and M. Ueda , Self-organization of the phosphatidylinositol lipids signaling system for random cell migration , Proc. Natl. Acad. Sci., 107 (2010), pp. 12399–12404
work page 2010
-
[4]
M. A. Avila Ponce de Le´on, B. F´elix, and H. G. Othmer, A phosphoinositide-based model of actin waves in frustrated phagocytosis , J. Theoret. Biol., 527 (2021), 110764. BISTABILITY OF WA VES AND WA VE-PINNING STATES 43 0 6.5 0 λ u v F (a) Q s≈ 0.500 0 6.5 0 3λ u v F (d) Q s≈ 0.542 0 6.5 0 3λ u v F (b) Q s≈ 0.500 −15 0 15 0 3λ (e)Re(F~) s≈ 0.542 0 6.5 0 ...
work page 2021
-
[5]
A. Bailles, E. W. Gehrels, and T. Lecuit , Mechanochemical principles of spatial and temporal patterns in cells and tissues , Annu. Rev. Cell Dev. Biol., 38 (2022), pp. 321–347
work page 2022
-
[6]
E. L. Barnhart, J. Allard, S. S. Lou, J. A. Theriot, and A. Mogilner , Adhesion-dependent wave generation in crawling cells , Curr. Biol., 27 (2017), pp. 27–38. 44 HUGHES, MODAI, EDELSTEIN-KESHET, AND YOCHELIS
work page 2017
-
[7]
W. M. Bement, A. B. Goryachev, A. L. Miller, and G. von Dassow, Patterning of the cell cortex by Rho GTPases , Nat. Rev. Mol. Cell Biol., 25 (2024), pp. 290–308
work page 2024
-
[8]
W. M. Bement, M. Leda, A. M. Moe, A. M. Kita, M. E. Larson, A. E. Golding, C. Pfeuti, K.-C. Su, A. L. Miller, A. B. Goryachev, and G. von Dassow , Activator–inhibitor coupling between Rho signalling and actin assembly makes the cell cortex an excitable medium , Nat. Cell Biol., 17 (2015), pp. 1471–1483
work page 2015
-
[9]
F. Bergmann, L. Rapp, and W. Zimmermann , Active phase separation: A universal approach , Phys. Rev. E, 98 (2018), 020603
work page 2018
-
[10]
F. Bergmann and W. Zimmermann, On system-spanning demixing properties of cell polarization, PloS One, 14 (2019), e0218328
work page 2019
-
[11]
E. Bernitt, H.-G. D ¨obereiner, N. S. Gov, and A. Yochelis , Fronts and waves of actin polymer- ization in a bistability-based mechanism of circular dorsal ruffles , Nat. Commun., 8 (2017), 15863
work page 2017
-
[12]
C. Beta, L. Edelstein-Keshet, N. Gov, and A. Yochelis , From actin waves to mechanism and back: How theory aids biological understanding , eLife, 12 (2023), e87181
work page 2023
-
[13]
C. Beta, N. S. Gov, and A. Yochelis , Why a large–scale mode can be essential for understanding intracellular actin waves , Cells, 9 (2020), 1533
work page 2020
-
[14]
C. Beta and K. Kruse , Intracellular oscillations and waves , Annu. Rev. Condens. Matter Phys., 8 (2017), pp. 239–264
work page 2017
-
[15]
W.-J. Beyn and V. Th ¨ummler, Phase conditions, symmetries and PDE continuation , in Numeri- cal Continuation Methods for Dynamical Systems: Path following and boundary value problems, B. Krauskopf, H. M. Osinga, and J. Gal´ an-Vioque, eds., Springer Netherlands, Dordrecht, 2007, pp. 301–330
work page 2007
- [16]
-
[17]
F. Brauns and M. C. Marchetti , Nonreciprocal pattern formation of conserved fields, Phys. Rev. X, 14 (2024), 021014
work page 2024
- [18]
-
[19]
B. A. Camley, Y. Zhao, B. Li, H. Levine, and W.-J. Rappel , Crawling and turning in a minimal reaction-diffusion cell motility model: Coupling cell shape and biochemistry , Phys. Rev. E, 95 (2017), 012401
work page 2017
-
[20]
Y. Cao, E. Ghabache, and W.-J. Rappel, Plasticity of cell migration resulting from mechanochemical coupling, eLife, 8 (2019), e48478
work page 2019
-
[21]
A. R. Champneys, F. Al Saadi, V. F. Bre ˜na-Medina, V. A. Grieneisen, A. F. Mar ´ee, N. Ver- schueren, and B. Wuyts , Bistability, wave pinning and localisation in natural reaction–diffusion systems, Phys D, 416 (2021), 132735
work page 2021
-
[22]
A. R. Champneys, V. Kirk, E. Knobloch, B. E. Oldeman, and J. Sneyd , When Shil’nikov meets Hopf in excitable systems , SIAM J. Appl. Dyn. Syst., 6 (2007), pp. 663–693
work page 2007
-
[23]
M. C. Cross and P. C. Hohenberg , Pattern formation outside of equilibrium , Rev. Modern Phys., 65 (1993), pp. 851–1123
work page 1993
-
[24]
H.-G. D¨obereiner, B. J. Dubin-Thaler, J. M. Hofman, H. S. Xenias, T. N. Sims, G. Giannone, M. L. Dustin, C. H. Wiggins, and M. P. Sheetz, Lateral membrane waves constitute a universal dynamic pattern of motile cells , Phys. Rev. Lett., 97 (2006), 038102
work page 2006
-
[25]
E. J. Doedel, A. R. Champneys, T. Fairgrieve, Y. Kuznetsov, B. Oldeman, R. Paffenroth, B. Sandstede, X. Wang, and C. Zhang , Auto07p: Continuation and bifurcation software for ordinary differential equations, 2012
work page 2012
-
[26]
K. Doubrovinski and K. Kruse, Cell motility resulting from spontaneous polymerization waves, Phys. Rev. Lett., 107 (2011), 258103
work page 2011
- [27]
-
[28]
C. M. Elliott, M. A. Herrero, J. R. King, and J. R. Ockendon , The mesa problem: Diffusion patterns for ut = ∇ · (um∇u) as m → +∞, IMA J. Appl. Math., 37 (1986), pp. 147–154
work page 1986
-
[29]
S. Fauve and O. Thual , Solitary waves generated by subcritical instabilities in dissipative systems , Phys. Rev. Lett., 64 (1990), 282. BISTABILITY OF WA VES AND WA VE-PINNING STATES 45
work page 1990
-
[30]
FitzHugh, Impulses and physiological states in theoretical models of nerve membrane , Biophys
R. FitzHugh, Impulses and physiological states in theoretical models of nerve membrane , Biophys. J., 1 (1961), pp. 445–466
work page 1961
-
[31]
G. Gerisch, M. Ecke, B. Schroth-Diez, S. Gerwig, U. Engel, L. Maddera, and M. Clarke , Self-organizing actin waves as planar phagocytic cup structures , Cell Adh. Migr., 3 (2009), pp. 373– 382
work page 2009
-
[32]
G. Giannone, B. J. Dubin-Thaler, H.-G. D ¨obereiner, N. Kieffer, A. R. Bresnick, and M. P. Sheetz, Periodic lamellipodial contractions correlate with rearward actin waves , Cell, 116 (2004), pp. 431–443
work page 2004
-
[33]
P. Glendinning and C. Sparrow, T-points: A codimension two heteroclinic bifurcation, J. Stat. Phys., 43 (1986), pp. 479––488
work page 1986
-
[34]
A. B. Goryachev and M. Leda, Many roads to symmetry breaking: Molecular mechanisms and theo- retical models of yeast cell polarity , Mol. Biol. Cell., 28 (2017), pp. 370–380
work page 2017
-
[35]
A. B. Goryachev, M. Leda, A. L. Miller, G. von Dassow, and W. M. Bement , How to make a static cytokinetic furrow out of traveling excitable waves , Small GTPases, 7 (2016), pp. 65–70
work page 2016
-
[36]
P. Hirschberg and E. Knobloch , ˇSil’nikov-hopf bifurcation, Phys. D, 62 (1993), pp. 202–216
work page 1993
-
[37]
S. Hladyshau, M. Kho, S. Nie, and D. Tsygankov , Spatiotemporal development of coexisting wave domains of rho activity in the cell cortex , Sci. Rep., 11 (2021), 19512
work page 2021
-
[38]
W. R. Holmes, A. E. Carlsson, and L. Edelstein-Keshet , Regimes of wave type patterning driven by refractory actin feedback: Transition from static polarization to dynamic wave behaviour , Phys. Biol., 9 (2012), 046005
work page 2012
-
[39]
W. R. Holmes, J. Park, A. Levchenko, and L. Edelstein-Keshet, A mathematical model coupling polarity signaling to cell adhesion explains diverse cell migration patterns , PLoS Comput. Biol., 13 (2017), e1005524
work page 2017
-
[40]
J. M. Hughes, C. Martinez-Torres, C. Beta, L. Edelstein-Keshet, and A. Yochelis , A dis- sipative mass conserved reaction–diffusion system reveals switching between coexisting polar and oscillatory cell motility states , Chaos, 35 (2025), 051103
work page 2025
-
[41]
N. Inagaki and H. Katsuno , Actin waves: Origin of cell polarization and migration? , Trends Cell Biol., 27 (2017), pp. 515–526
work page 2017
- [42]
-
[43]
A. Jilkine and L. Edelstein-Keshet, A comparison of mathematical models for polarization of single eukaryotic cells in response to guided cues , PLoS Comput. Biol., 7 (2011), e1001121
work page 2011
-
[44]
E. Knobloch, On the degenerate Hopf bifurcation with O(2) symmetry , in Multiparameter bifurcation theory, M. Golubitsky and J. Guckenheimer, eds., vol. 56 of Contemporary Mathematics, American Mathematical Society, 1986, pp. 193—-201
work page 1986
-
[45]
Knobloch, Oscillatory convection in binary mixtures , Phys
E. Knobloch, Oscillatory convection in binary mixtures , Phys. Rev. A, 34 (1986), 1538
work page 1986
-
[46]
Knobloch, Spatial localization in dissipative systems , Annu
E. Knobloch, Spatial localization in dissipative systems , Annu. Rev. Condens. Matter Phys., 6 (2015), pp. 325–359
work page 2015
-
[47]
E. Knobloch and A. Yochelis, Front propagation and global bifurcations in a multivariable reaction- diffusion model, Chaos, 33 (2023)
work page 2023
-
[48]
T. Kolokolnikov, T. Erneux, and J. Wei , Mesa-type patterns in the one-dimensional brusselator and their stability , Phys. D, 214 (2006), pp. 63–77
work page 2006
-
[49]
M. Kuwamura, T. Teramoto, and H. Ikeda , Single transition layer in mass-conserving reaction- diffusion systems with bistable nonlinearity , Nonlinearity, 37 (2024), 115013
work page 2024
-
[50]
Y. A. Kuznetsov, Elements of applied bifurcation theory , Springer, 2004
work page 2004
-
[51]
J. Landino, M. Leda, A. Michaud, Z. T. Swider, M. Prom, C. M. Field, W. M. Bement, A. G. Vecchiarelli, A. B. Goryachev, and A. L. Miller , Rho and F-actin self-organize within an artificial cell cortex, Curr. Biol., 31 (2021), pp. 5613–5621
work page 2021
-
[52]
Z. D. Li, C. Liu, A. van Kan, and E. Knobloch , Traveling spatially localized convective structures in an inclined porous medium , Phys. Rev. Fluids, 10 (2025), 034402
work page 2025
-
[53]
Y. Liu, E. G. Rens, and L. Edelstein-Keshet , Spots, stripes, and spiral waves in models for static and motile cells , J. Math. Biol., 82 (2021), pp. 1–38
work page 2021
-
[54]
M. Machacek and G. Danuser , Morphodynamic profiling of protrusion phenotypes , Biophys. J., 90 (2006), pp. 1439–1452. 46 HUGHES, MODAI, EDELSTEIN-KESHET, AND YOCHELIS
work page 2006
-
[55]
A. F. Mar ´ee, V. A. Grieneisen, and L. Edelstein-Keshet , How cells integrate complex stimuli: The effect of feedback from phosphoinositides and cell shape on cell polarization and motility , PLoS Comput. Biol., 8 (2012), e1002402
work page 2012
-
[56]
P. C. Matthews and S. M. Cox, Pattern formation with a conservation law , Nonlinearity, 13 (2000), pp. 1293––1320
work page 2000
-
[57]
A. Michaud, M. Leda, Z. T. Swider, S. Kim, J. He, J. Landino, J. R. Valley, J. Huisken, A. B. Goryachev, G. von Dassow, et al. , A versatile cortical pattern-forming circuit based on Rho, F-actin, Ect2 and RGA-3/4 , J. Cell Biol., 221 (2022), e202203017
work page 2022
-
[58]
A. Michaud, Z. T. Swider, J. Landino, M. Leda, A. L. Miller, G. von Dassow, A. B. Gory- achev, and W. M. Bement, Cortical excitability and cell division, Curr. Biol., 31 (2021), pp. R553– R559
work page 2021
-
[59]
P. W. Miller, D. Fortunato, M. Novaga, S. Y. Shvartsman, and C. B. Muratov , Generation and motion of interfaces in a mass-conserving reaction-diffusion system , SIAM J. Appl. Dyn. Syst., 22 (2023), pp. 2408–2431
work page 2023
-
[60]
T. Moldenhawer, E. Moreno, D. Schindler, S. Flemming, M. Holschneider, W. Huisinga, S. Alonso, and C. Beta , Spontaneous transitions between amoeboid and keratocyte-like modes of migration, Front. Cell Dev. Biol., 10 (2022), 898351
work page 2022
- [61]
-
[62]
P. Moreno-Spiegelberg, A. Arinyo-i Prats, D. Ruiz-Reyn ´es, M. A. Matias, and D. Gomila , Bifurcation structure of traveling pulses in type-i excitable media, Phys. Rev. E, 106 (2022), p. 034206
work page 2022
-
[63]
Y. Mori, A. Jilkine, and L. Edelstein-Keshet , Wave-pinning and cell polarity from a bistable reaction-diffusion system, Biophys. J., 94 (2008), pp. 3684–3697
work page 2008
-
[64]
Y. Mori, A. Jilkine, and L. Edelstein-Keshet, Asymptotic and bifurcation analysis of wave-pinning in a reaction-diffusion model for cell polarization , SIAM J. Appl. Math., 71 (2011), pp. 1401–1427
work page 2011
-
[65]
J. D. Murray , Mathematical Biology II: Spatial models and biomedical applications , Springer, New York, 2003
work page 2003
-
[66]
J. D. Murray, Mathematical Biology: I. An Introduction , vol. 17, Springer Science & Business Media, 2007
work page 2007
- [67]
-
[68]
M. Or-Guil, J. Krishnan, I. Kevrekidis, and M. B ¨ar, Pulse bifurcations and instabilities in an excitable medium: Computations in finite ring domains , Phys. Rev. E, 64 (2001), 046212
work page 2001
- [69]
-
[70]
P. Parra-Rivas, E. Knobloch, D. Gomila, and L. Gelens , Dark solitons in the Lugiato-Lefever equation with normal dispersion , Phys. Rev. A, 93 (2016), 063839
work page 2016
-
[71]
C. Rackauckas and Q. Nie, Differentialequations.jl–a performant and feature-rich ecosystem for solv- ing differential equations in Julia , J. Open Res. Softw., 5 (2017), 15
work page 2017
-
[72]
M. Raja, A. van Kan, B. Foster, and E. Knobloch , Collisions of localized patterns in a nonvari- ational swift-hohenberg equation, Phys. Rev. E, 107 (2023), 064214
work page 2023
-
[73]
E. G. Rens and L. Edelstein-Keshet, Cellular tango: How extracellular matrix adhesion choreographs rac-rho signaling and cell movement , Phys. Biol., 18 (2021), 066005
work page 2021
-
[74]
A. J. Ridley, Rho GTPase signalling in cell migration , Curr. Opin. Cell Biol., 36 (2015), pp. 103–112
work page 2015
-
[75]
J. F. Robinson, T. Machon, and T. Speck, Universal limiting behavior of reaction-diffusion systems with conservation laws , Phys. Rev. E, 111 (2025), 065417
work page 2025
-
[76]
M. M. Romeo and C. K. Jones , The stability of traveling calcium pulses in a pancreatic acinar cell , Phys. D, 177 (2003), pp. 242–258
work page 2003
-
[77]
J. A. Sherratt, M. A. Lewis, and A. C. Fowler , Ecological chaos in the wake of invasion. , Proc. Natl. Acad. Sci., 92 (1995), pp. 2524—-2528
work page 1995
-
[78]
J. A. Sherratt, M. J. Smith, and J. D. Rademacher , Locating the transition from periodic oscilla- tions to spatiotemporal chaos in the wake of invasion , Proc. Natl. Acad. Sci., 106 (2009), pp. 10890– 10895. BISTABILITY OF WA VES AND WA VE-PINNING STATES 47
work page 2009
- [79]
-
[80]
G. Steinebach , Construction of Rosenbrock–Wanner method Rodas5P and numerical benchmarks within the Julia differential equations package , BIT Numer. Math., 63 (2023), 27
work page 2023
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.