Symmetry-protected phases in a 1D active solid with mechanochemical feedback
Pith reviewed 2026-05-22 19:06 UTC · model grok-4.3
The pith
Reciprocal coupling of elasticity and Hopf oscillators in a 1D active solid produces symmetry-protected phases and a universal transition to compression-driven oscillation death.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
Complex self-organization in active solids with mechanochemical feedback can be classified purely through symmetry arguments, yielding a rich landscape of symmetry-protected phases together with a universal transition to compression-driven oscillation death that supplies a physical account of localized signaling dampening in biological tissues.
What carries the argument
Reciprocal coupling between elasticity and Hopf oscillators, analyzed through amplitude equations and group-theoretic symmetry classification.
If this is right
- Symmetry arguments alone suffice to predict the full set of stable phases in such active solids.
- The compression-driven oscillation death transition supplies a mechanism for localized dampening of chemical signals inside tissues.
- Inconsistencies in earlier models of signaling dampening are resolved by the existence of this universal transition.
- Self-organized patterns in active solids follow universal symmetry rules independent of most microscopic details.
Where Pith is reading between the lines
- The same symmetry classification could be extended to two- or three-dimensional active solids to predict additional protected states.
- Synthetic active materials could be engineered to exhibit controllable oscillation death for precise spatial signaling.
- Experiments that vary compression while tracking oscillation amplitude in real cell chains would directly test the predicted transition.
Load-bearing premise
The reciprocal coupling between elasticity and Hopf oscillators in the one-dimensional model captures the essential mechanochemical feedback present in real biological tissues.
What would settle it
Direct observation that compression applied to a one-dimensional chain of coupled oscillators fails to produce uniform oscillation death, or that the observed phases violate the symmetry-protected pattern predicted by the amplitude equations, would refute the central claim.
Figures



read the original abstract
We present a framework for mechanochemical self-organization in active solids where elasticity is reciprocally coupled to Hopf oscillators. Our model reveals a rich landscape of symmetry-protected phases, identified through amplitude equations and group-theoretic analysis. We uncover a universal transition to compression-driven oscillation death (COD), providing a physical basis for localized signaling dampening in biological tissues that resolves inconsistencies in previous models. Our work demonstrates that complex self-organization in active solids can be classified purely through symmetry arguments.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript develops a 1D active solid model in which elasticity is reciprocally coupled to a lattice of Hopf oscillators. Amplitude equations are derived from this coupling and combined with group-theoretic analysis to classify a landscape of symmetry-protected phases; a universal compression-driven oscillation death (COD) transition is identified and proposed as a mechanism for localized signaling dampening in biological tissues.
Significance. If the amplitude-equation reduction remains valid through the COD point and the symmetry classification is free of hidden parameter dependence, the work supplies a symmetry-based taxonomy for self-organization in active solids that could unify disparate observations in mechanochemical systems. The explicit link to tissue-level signaling dampening is a concrete biophysical implication.
major comments (2)
- [Amplitude equations and COD transition] The derivation of the amplitude equations (presumably in the section following the model definition) assumes that the oscillators remain in the limit-cycle regime with slowly varying amplitudes up to the COD transition. In a 1D chain under increasing compression, local amplitude can reach zero before the slow-variation assumption holds, converting the purported symmetry-protected COD into an ordinary supercritical Hopf bifurcation. An explicit center-manifold or multiple-scale validity check at the transition is required to substantiate the universality claim.
- [Group-theoretic analysis] The group-theoretic classification of phases relies on the reciprocal elasticity-Hopf coupling preserving the requisite symmetries. The manuscript should demonstrate that no effective parameters are introduced by the coupling that would break the symmetry protection or render the phase diagram non-universal (cf. the claim that classification is 'purely through symmetry arguments').
minor comments (1)
- [Abstract] The abstract states that the COD transition 'resolves inconsistencies in previous models' without naming the models or the inconsistencies; a single sentence identifying them would improve readability.
Simulated Author's Rebuttal
We thank the referee for the careful reading and constructive comments on our manuscript. We address each major point below and indicate the revisions that will be made to strengthen the presentation of the amplitude-equation validity and the symmetry arguments.
read point-by-point responses
-
Referee: [Amplitude equations and COD transition] The derivation of the amplitude equations (presumably in the section following the model definition) assumes that the oscillators remain in the limit-cycle regime with slowly varying amplitudes up to the COD transition. In a 1D chain under increasing compression, local amplitude can reach zero before the slow-variation assumption holds, converting the purported symmetry-protected COD into an ordinary supercritical Hopf bifurcation. An explicit center-manifold or multiple-scale validity check at the transition is required to substantiate the universality claim.
Authors: We agree that an explicit validity check is required. In the revised manuscript we will add a center-manifold analysis together with direct numerical integration of the full oscillator-elasticity system. These checks confirm that, for the adiabatic compression rates used, the slow-amplitude assumption remains valid through the COD point and that the transition retains its symmetry-protected character rather than reducing to a standard supercritical Hopf bifurcation. revision: yes
-
Referee: [Group-theoretic analysis] The group-theoretic classification of phases relies on the reciprocal elasticity-Hopf coupling preserving the requisite symmetries. The manuscript should demonstrate that no effective parameters are introduced by the coupling that would break the symmetry protection or render the phase diagram non-universal (cf. the claim that classification is 'purely through symmetry arguments').
Authors: The reciprocal coupling is constructed to be invariant under the full symmetry group of the one-dimensional chain (translations and reflections). In the revised manuscript we will insert an explicit symmetry-transformation table for the coupling terms, demonstrating that no additional parameters appear that break these symmetries. Consequently the phase classification remains universal and rests solely on group-theoretic arguments. revision: yes
Circularity Check
No significant circularity; derivation self-contained in symmetry and amplitude analysis
full rationale
The provided abstract and context describe a model coupling elasticity to Hopf oscillators, deriving amplitude equations, and applying group theory to classify phases and identify a COD transition. No equations, parameter fits, or self-citations are quoted that reduce any prediction or phase classification to an input by construction. The central claims rest on the reciprocal coupling framework and symmetry arguments presented as derived outputs rather than redefined inputs. Absent explicit text showing a fitted coupling renamed as a universal prediction or a self-citation chain justifying uniqueness, the derivation does not exhibit the enumerated circular patterns. This is the expected honest non-finding for a symmetry-based classification paper whose details are not shown to collapse into tautology.
Axiom & Free-Parameter Ledger
Lean theorems connected to this paper
-
IndisputableMonolith/Foundation/AbsoluteFloorClosure.leanreality_from_one_distinction unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
Our model reveals a rich landscape of symmetry-protected phases, identified through amplitude equations and group-theoretic analysis. We uncover a universal transition to compression-driven oscillation death (COD)
-
IndisputableMonolith/Foundation/AlexanderDuality.leanalexander_duality_circle_linking unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
the underlying symmetry of HBR implies that the observed patterns and phases may generically arise in many natural and synthetic systems
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
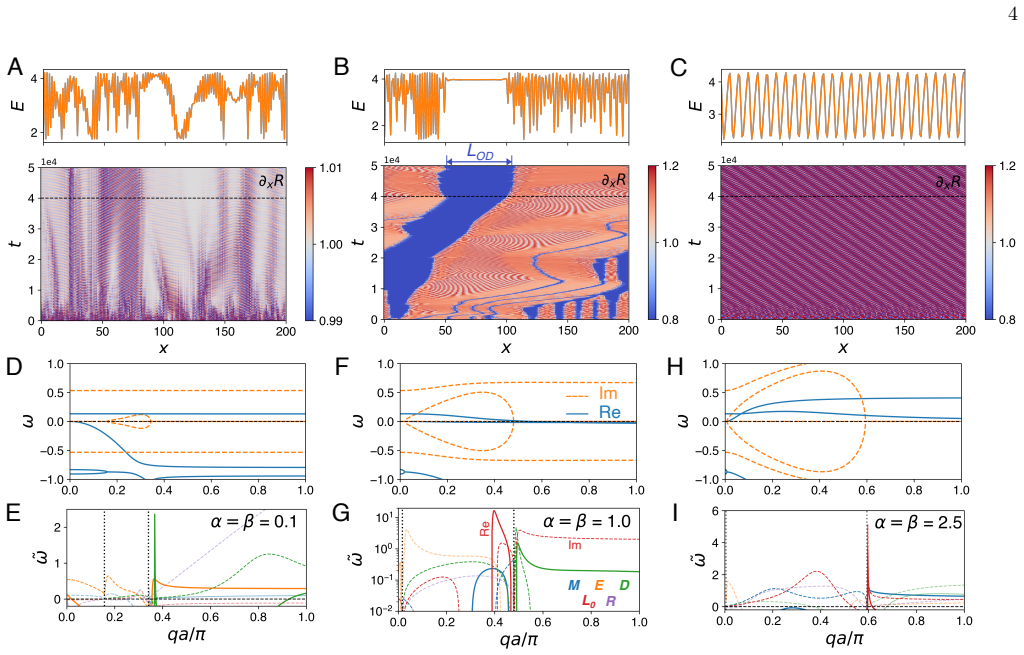
-
[1]
W. Lim, B. Mayer, and T. Pawson, Cell signaling (Gar- land Science, 2014)
work page 2014
- [2]
-
[3]
E. Hannezo and C.-P. Heisenberg, Mechanochemical Feedback Loops in Development and Disease, Cell 178, 12 (2019), publisher: Elsevier
work page 2019
-
[4]
D. B. Br¨ uckner and E. Hannezo, Tissue Active Mat- ter: Integrating Mechanics and Signaling into Dynam- ical Models, Cold Spring Harbor Perspectives in Biology , a041653 (2024), company: Cold Spring Harbor Labora- tory Press Distributor: Cold Spring Harbor Laboratory Press Institution: Cold Spring Harbor Laboratory Press Label: Cold Spring Harbor Laborat...
work page 2024
-
[5]
A. Ioratim-Uba, T. B. Liverpool, and S. Henkes, Mechanochemical Active Feedback Generates Conver- gence Extension in Epithelial Tissue, Physical Review Letters 131, 238301 (2023)
work page 2023
-
[6]
H. Lan, Q. Wang, R. Fernandez-Gonzalez, and J. J. Feng, A biomechanical model for cell polarization and interca- lation during drosophila germband extension, Physical biology 12, 056011 (2015)
work page 2015
-
[7]
S. Banerjee, K. J. Utuje, and M. C. Marchetti, Propa- gating stress waves during epithelial expansion, Physical review letters 114, 228101 (2015)
work page 2015
-
[8]
R. Sknepnek, I. Djafer-Cherif, M. Chuai, C. Weijer, and S. Henkes, Generating active t1 transitions through mechanochemical feedback, Elife 12, e79862 (2023)
work page 2023
-
[9]
K. Dierkes, A. Sumi, J. Solon, and G. Salbreux, Spon- taneous oscillations of elastic contractile materials with turnover, Physical review letters 113, 148102 (2014)
work page 2014
-
[10]
M. F. Staddon, K. E. Cavanaugh, E. M. Munro, M. L. Gardel, and S. Banerjee, Mechanosensitive junction re- modeling promotes robust epithelial morphogenesis, Bio- physical Journal 117, 1739 (2019)
work page 2019
-
[11]
K. E. Cavanaugh, M. F. Staddon, S. Banerjee, and M. L. Gardel, Adaptive viscoelasticity of epithelial cell junc- tions: from models to methods, Current opinion in ge- netics & development 63, 86 (2020)
work page 2020
-
[12]
R. Etournay, M. Popovi´ c, M. Merkel, A. Nandi, C. Blasse, B. Aigouy, H. Brandl, G. Myers, G. Salbreux, F. J¨ ulicher,et al. , Interplay of cell dynamics and epithe- lial tension during morphogenesis of the drosophila pupal wing, Elife 4, e07090 (2015)
work page 2015
-
[13]
N. Noll, M. Mani, I. Heemskerk, S. J. Streichan, and B. I. Shraiman, Active tension network model suggests an exotic mechanical state realized in epithelial tissues, Nature physics 13, 1221 (2017)
work page 2017
-
[14]
T. B. Saw, A. Doostmohammadi, V. Nier, L. Kocgozlu, S. Thampi, Y. Toyama, P. Marcq, C. T. Lim, J. M. Yeo- mans, and B. Ladoux, Topological defects in epithelia govern cell death and extrusion, Nature 544, 212 (2017)
work page 2017
-
[15]
S. J. Streichan, M. F. Lefebvre, N. Noll, E. F. Wieschaus, and B. I. Shraiman, Global morphogenetic flow is accu- rately predicted by the spatial distribution of myosin mo- tors, Elife 7, e27454 (2018)
work page 2018
-
[16]
M. Ibrahimi and M. Merkel, Deforming polar active mat- ter in a scalar field gradient, New Journal of Physics 25, 013022 (2023)
work page 2023
-
[17]
R. Cl´ ement, B. Dehapiot, C. Collinet, T. Lecuit, and P.-F. Lenne, Viscoelastic dissipation stabilizes cell shape changes during tissue morphogenesis, Current biology 27, 3132 (2017)
work page 2017
-
[18]
N. Khalilgharibi, J. Fouchard, N. Asadipour, R. Bar- rientos, M. Duda, A. Bonfanti, A. Yonis, A. Harris, P. Mosaffa, Y. Fujita, et al. , Stress relaxation in epithe- lial monolayers is controlled by the actomyosin cortex, Nature physics 15, 839 (2019)
work page 2019
-
[19]
L. W¨ urthner, A. Goychuk, and E. Frey, Geometry- induced patterns through mechanochemical coupling, Physical Review E 108, 014404 (2023)
work page 2023
-
[20]
T. Banerjee, T. Desaleux, J. Ranft, and ´E. Fodor, Hy- drodynamics of pulsating active liquids, arXiv preprint arXiv:2407.19955 (2024)
-
[21]
Mathematical modelling of calcium signalling taking into account mechanical effects
K. Kaouri, P. K. Maini, and S. J. Chapman, Mathemati- cal modelling of calcium signalling taking into account mechanical effects, arXiv preprint arXiv:1703.00540 (2017). 7
work page internal anchor Pith review Pith/arXiv arXiv 2017
-
[22]
Effect of curvature and normal forces on motor regulation of cilia
P. Sartori, Effect of curvature and normal forces on mo- tor regulation of cilia, arXiv preprint arXiv:1905.04138 (2019)
work page internal anchor Pith review Pith/arXiv arXiv 1905
- [23]
-
[24]
A. Dhanuka, D. S. Banerjee, M. Rao, et al. , Excitability and travelling waves in renewable active matter, arXiv preprint arXiv:2503.19687 (2025)
-
[25]
H. De Belly, E. K. Paluch, and K. J. Chalut, Interplay between mechanics and signalling in regulating cell fate, Nature Reviews Molecular Cell Biology 23, 465 (2022)
work page 2022
-
[26]
D. Boocock, Interplay between Mechanochemical Pat- terning and Glassy Dynamics in Cellular Monolayers, PRX Life 1, 10.1103/PRXLife.1.013001 (2023)
-
[27]
D. Boocock, N. Hino, N. Ruzickova, T. Hirashima, and E. Hannezo, Theory of mechanochemical patterning and optimal migration in cell monolayers, Nature Physics 17, 267 (2021), publisher: Nature Publishing Group
work page 2021
-
[28]
F. P´ erez-Verdugo, S. Banks, and S. Banerjee, Excitable dynamics driven by mechanical feedback in biological tissues, Communications Physics 7, 1 (2024), publisher: Nature Publishing Group
work page 2024
- [29]
-
[30]
H. Shankaran, D. L. Ippolito, W. B. Chrisler, H. Resat, N. Bollinger, L. K. Opresko, and H. S. Wiley, Rapid and sustained nuclear–cytoplasmic ERK oscillations induced by epidermal growth factor, Molecular Systems Biology 5, 332 (2009)
work page 2009
- [31]
-
[32]
N. Hino, L. Rossetti, A. Mar´ ın-Llaurad´ o, K. Aoki, X. Trepat, M. Matsuda, and T. Hirashima, Erk-mediated mechanochemical waves direct collective cell polariza- tion, Developmental cell 53, 646 (2020)
work page 2020
-
[33]
J. C. Barros and C. J. Marshall, Activation of either erk1/2 or erk5 map kinase pathways can lead to disrup- tion of the actin cytoskeleton, Journal of cell science118, 1663 (2005)
work page 2005
-
[34]
J. S. Logue, A. X. Cartagena-Rivera, M. A. Baird, M. W. Davidson, R. S. Chadwick, and C. M. Waterman, Erk regulation of actin capping and bundling by eps8 pro- motes cortex tension and leader bleb-based migration, elife 4, e08314 (2015)
work page 2015
- [35]
-
[36]
M. C. Mendoza, E. E. Er, W. Zhang, B. A. Ballif, H. L. Elliott, G. Danuser, and J. Blenis, Erk-mapk drives lamellipodia protrusion by activating the wave2 regula- tory complex, Molecular cell 41, 661 (2011)
work page 2011
-
[37]
M. C. Mendoza, M. Vilela, J. E. Juarez, J. Blenis, and G. Danuser, Erk reinforces actin polymerization to power persistent edge protrusion during motility, Science signal- ing 8, ra47 (2015)
work page 2015
-
[38]
S. Tanimura and K. Takeda, Erk signalling as a regulator of cell motility, The Journal of Biochemistry 162, 145 (2017)
work page 2017
-
[39]
H. De Belly, A. Stubb, A. Yanagida, C. Labouesse, P. H. Jones, E. K. Paluch, and K. J. Chalut, Membrane tension gates erk-mediated regulation of pluripotent cell fate, Cell stem cell 28, 273 (2021)
work page 2021
-
[40]
B. N. Kholodenko, J. F. Hancock, and W. Kolch, Sig- nalling ballet in space and time, Nature reviews Molecu- lar cell biology 11, 414 (2010)
work page 2010
-
[41]
Y. Arkun and M. Yasemi, Dynamics and control of the erk signaling pathway: Sensitivity, bistability, and oscil- lations, PloS one 13, e0195513 (2018)
work page 2018
-
[42]
K. Aoki, Y. Kondo, H. Naoki, T. Hiratsuka, R. E. Itoh, and M. Matsuda, Propagating wave of erk activation ori- ents collective cell migration, Developmental cell 43, 305 (2017)
work page 2017
-
[43]
J. J. Tyson, Biochemical Oscillations, in Computational Cell Biology , Vol. 20, edited by C. P. Fall, E. S. Marland, J. M. Wagner, and J. J. Tyson (Springer New York, New York, NY, 2004) pp. 230–260, series Title: Interdisci- plinary Applied Mathematics
work page 2004
-
[44]
I. R. Epstein and J. A. Pojman, An introduction to non- linear chemical dynamics: oscillations, waves, patterns, and chaos (Oxford university press, 1998)
work page 1998
-
[45]
J. J. Collins and I. N. Stewart, Coupled nonlinear os- cillators and the symmetries of animal gaits, Journal of Nonlinear Science 3, 349 (1993)
work page 1993
-
[46]
J. J. Collins and I. Stewart, A group-theoretic approach to rings of coupled biological oscillators, Biological Cy- bernetics 71, 95 (1994)
work page 1994
-
[47]
I. R. Epstein and M. Golubitsky, Symmetric patterns in linear arrays of coupled cells, Chaos: An Interdisciplinary Journal of Nonlinear Science 3, 1 (1993)
work page 1993
-
[48]
International Centre for Theoretical Sciences, Dynamical phase transitions in Markov processes by Hugo Touchette (2019)
work page 2019
-
[49]
S. H. Strogatz, Nonlinear dynamics and chaos: with ap- plications to physics, biology, chemistry, and engineering (Chapman and Hall/CRC, 2024)
work page 2024
-
[50]
T. Mora and W. Bialek, Are biological systems poised at criticality?, Journal of Statistical Physics 144, 268 (2011)
work page 2011
-
[51]
M. A. Munoz, Colloquium: Criticality and dynamical scaling in living systems, Reviews of Modern Physics 90, 031001 (2018)
work page 2018
- [52]
-
[53]
C. Scheibner, A. Souslov, D. Banerjee, P. Sur´ owka, W. T. Irvine, and V. Vitelli, Odd elasticity, Nature Physics 16, 475 (2020)
work page 2020
-
[54]
D. Banerjee, V. Vitelli, F. J¨ ulicher, and P. Sur´ owka, Ac- tive viscoelasticity of odd materials, Physical Review Let- ters 126, 138001 (2021)
work page 2021
-
[55]
A. Koseska, E. Volkov, and J. Kurths, Oscillation quench- ing mechanisms: Amplitude vs. oscillation death, Physics Reports 531, 173 (2013)
work page 2013
-
[56]
M. Fruchart, R. Hanai, P. B. Littlewood, and V. Vitelli, Non-reciprocal phase transitions, Nature 592, 363 (2021)
work page 2021
-
[57]
Davidson, Turbulence: an introduction for scientists and engineers (Oxford university press, 2015)
P. Davidson, Turbulence: an introduction for scientists and engineers (Oxford university press, 2015)
work page 2015
-
[58]
P. T. Nyawo and H. Touchette, A minimal model of dy- namical phase transition, Europhysics Letters116, 50009 (2017)
work page 2017
-
[59]
Wechselberger, Canards, Scholarpedia 2, 1356 (2007)
M. Wechselberger, Canards, Scholarpedia 2, 1356 (2007)
work page 2007
-
[60]
Coexistence of Coherence and Incoherence in Nonlocally Coupled Phase Oscillators
Y. Kuramoto and D. Battogtokh, Coexistence of coher- ence and incoherence in nonlocally coupled phase oscil- lators, arXiv preprint cond-mat/0210694 (2002). 8
work page internal anchor Pith review Pith/arXiv arXiv 2002
-
[61]
D. M. Abrams and S. H. Strogatz, Chimera states for coupled oscillators, Physical review letters 93, 174102 (2004)
work page 2004
-
[62]
E. A. Martens, S. Thutupalli, A. Fourri` ere, and O. Hal- latschek, Chimera states in mechanical oscillator net- works, Proceedings of the National Academy of Sciences 110, 10563 (2013)
work page 2013
-
[63]
J. Das, A. Chaudhuri, and S. Sinha, From order to chimeras: Unraveling dynamic patterns in active fluids with nonlinear growth, arXiv preprint arXiv:2412.14729 (2024)
work page internal anchor Pith review arXiv 2024
-
[64]
Sindhu M. et al. unpublished data
-
[65]
N. Roshan and R. Pandit, Multiscale studies of delayed afterdepolarizations i: A comparison of two biophysically realistic mathematical models for human ventricular my- ocytes, arXiv preprint arXiv:2307.10134 (2023)
-
[66]
N. Tompkins, N. Li, C. Girabawe, M. Heymann, G. B. Er- mentrout, I. R. Epstein, and S. Fraden, Testing turing’s theory of morphogenesis in chemical cells, Proceedings of the National Academy of Sciences 111, 4397 (2014)
work page 2014
- [67]
-
[68]
Y. Zhai, I. Z. Kiss, and J. L. Hudson, Amplitude death through a hopf bifurcation in coupled electrochemical os- cillators: Experiments and simulations, Physical Review E 69, 026208 (2004)
work page 2004
-
[69]
P. Baconnier, D. Shohat, C. H. L´ opez, C. Coulais, V. D´ emery, G. D¨ uring, and O. Dauchot, Selective and collective actuation in active solids, Nature Physics 18, 1234 (2022)
work page 2022
- [70]
-
[71]
D. C. Brody, Biorthogonal quantum mechanics, Journal of Physics A: Mathematical and Theoretical 47, 035305 (2013). 9 SUPPLEMENTARY INFORMATION: OSCILLATION DEATH BY MECHANOCHEMICAL FEEDBACK I. METHODS A. Why five variables? Our model solves the coupled dynamics of five variables: M, E, D, L0, R. L0 and R are the parameters of the spring and cannot be remo...
work page 2013
-
[72]
They are the unit of length and time in our model
a = 1 and τR = 1 for all simulations. They are the unit of length and time in our model
-
[73]
For Fig 3A-C, we have simulated a model with Lx = 200a and tmax = 5 × 104τR. A single seed value is shown
-
[74]
For Fig 4A-B, we have chosen α = β = √µ, where µ ∈ [1e − 2, 9]. Lx = 1000a, tmax = 105τR. The distributions were obtained from datapoints from t = 9.9 × 104 − 105 and averaged over 10 random initial conditions. All together, 103 × 103 × 10 = 107 samples were used to compute the distributions
-
[75]
S5, S6, we have chosen α, β = {0.1, 0.2, 0.3, 0.5, 0.8, 1, 1.5, 2, 2.5, 3}
For Fig 4C and Fig. S5, S6, we have chosen α, β = {0.1, 0.2, 0.3, 0.5, 0.8, 1, 1.5, 2, 2.5, 3}. For each of the 100 pairs of α, β values and 9 pairs of τD, τL values, we used 20 random initial conditions
- [76]
-
[77]
10 different replicates were used
For Fig.4E, we chose µ = µc ± δµ, where µc = 0.5, 3.24, and 5.60 and δµ ∈ [10−6, 10−1]. 10 different replicates were used. FIG. S2. Numerical estimate of LOD at short (left) and long timescales (right). The white lines show our estimate. The color map indicates the value of ∂xR. D. Numerical estimation of LOD We observed that E does not change appreciably...
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.