Enzyme-Substrate Complex Formation Modulates Diffusion-Driven Patterning In Metabolic Pathways
Pith reviewed 2026-05-17 01:13 UTC · model grok-4.3
The pith
Reversible enzyme-substrate binding modifies the conditions for diffusion-driven pattern formation in metabolic pathways.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
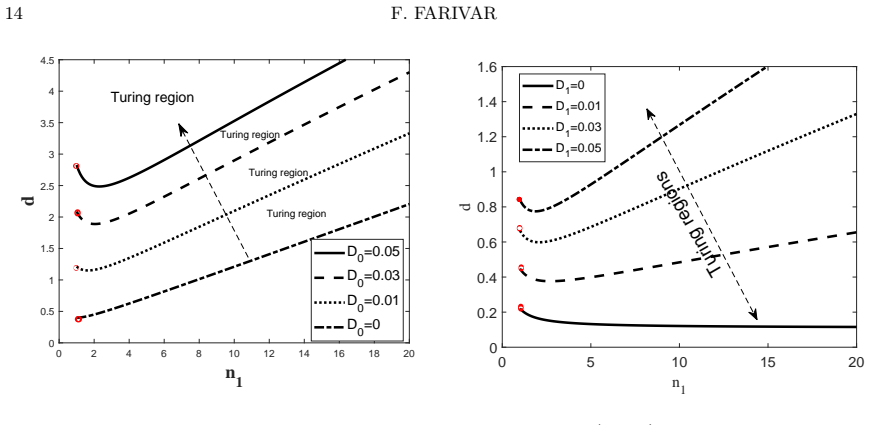
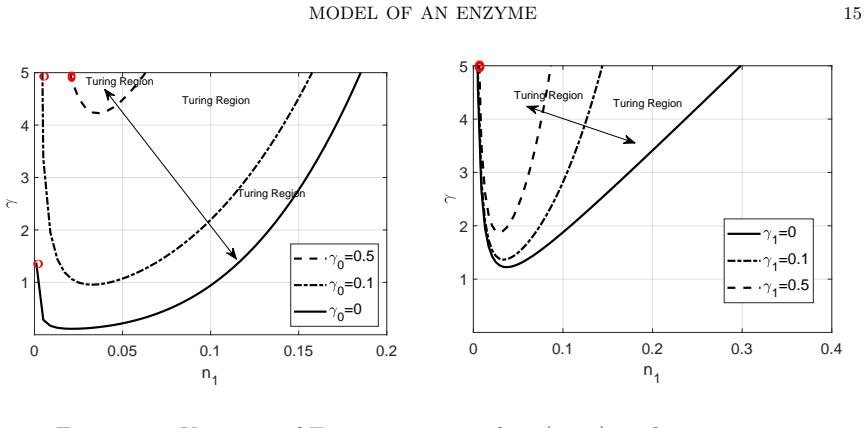
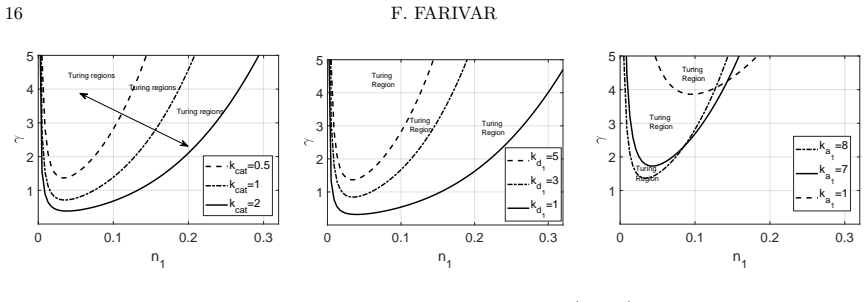
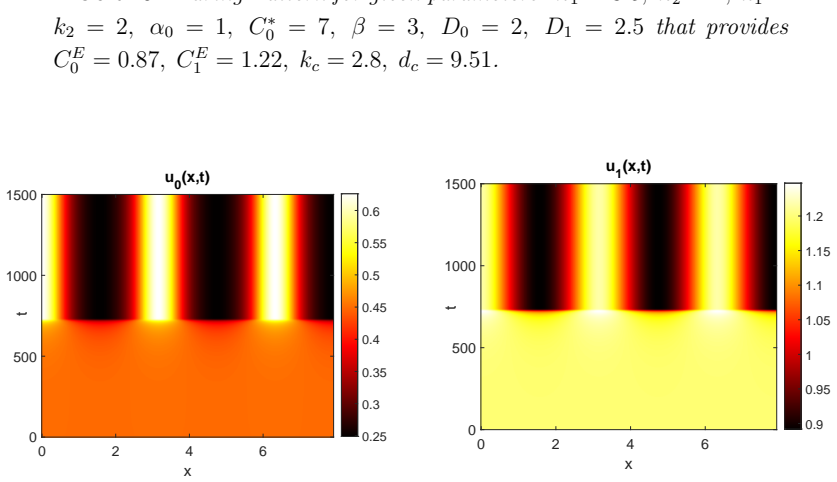
Starting from a mechanistic model that includes reversible enzyme-substrate complex formation, the analysis demonstrates that the quasi-steady-state reduced system exhibits a shifted Turing instability region compared to effective kinetics models. Weakly nonlinear analysis and simulations confirm that binding interactions alter pattern selection and slow the development of spatial heterogeneity in metabolite concentrations.
What carries the argument
The quasi-steady-state approximation applied to the enzyme-substrate complex, which eliminates the fast binding variable while retaining the nonlinear effects on the reaction terms.
If this is right
- The location and size of the parameter region allowing diffusion-driven instability depend on the binding and unbinding rates.
- Reversible binding can either promote or suppress pattern formation relative to irreversible or effective models.
- Pattern emergence is slower when enzyme-substrate interactions are accounted for explicitly.
- These changes provide a mechanistic explanation for mesoscale metabolic organization.
Where Pith is reading between the lines
- If binding kinetics vary across enzymes, different metabolic steps could have distinct pattern-forming tendencies.
- This framework could be extended to longer pathways to see how binding propagates spatial structure.
- Experimental tests might involve varying enzyme concentrations or using mutants with altered binding affinities to observe changes in metabolite clustering.
Load-bearing premise
The binding and unbinding of substrate to enzyme occur on a much faster time scale than changes in the overall metabolite concentrations.
What would settle it
A direct measurement of the critical diffusion coefficients or reaction rates at which patterns first appear in a controlled metabolic system that does not match the boundaries predicted by the stability analysis of the reduced model.
Figures

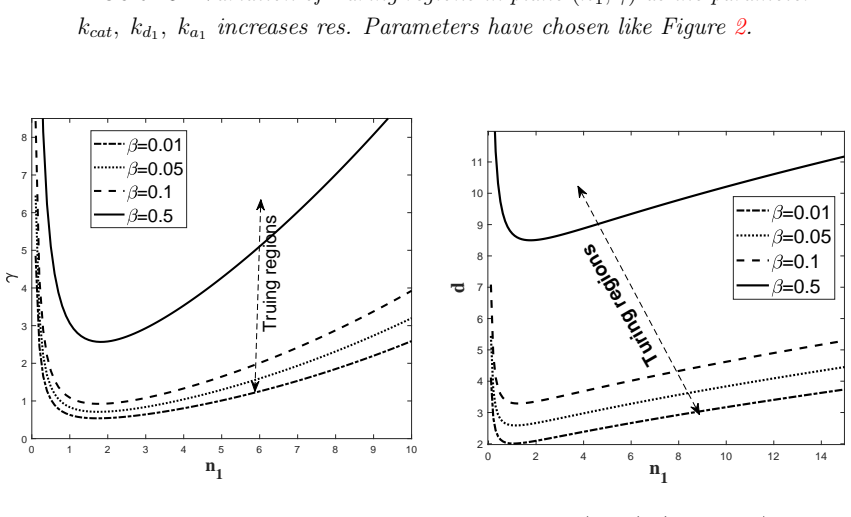
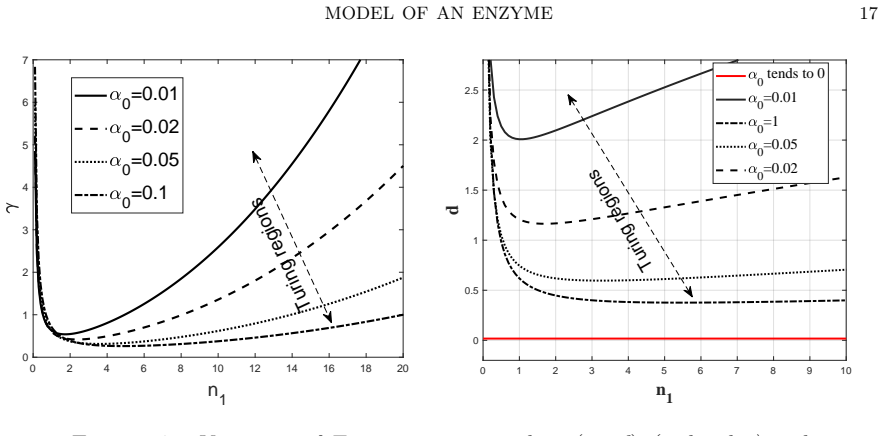
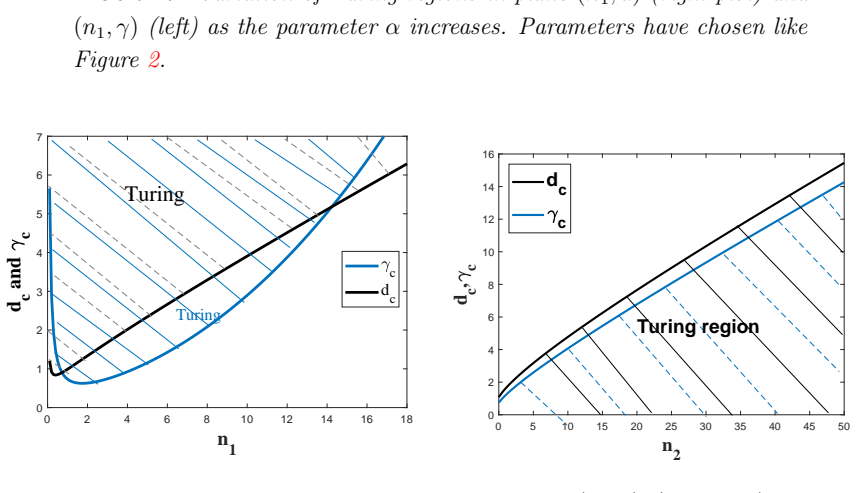
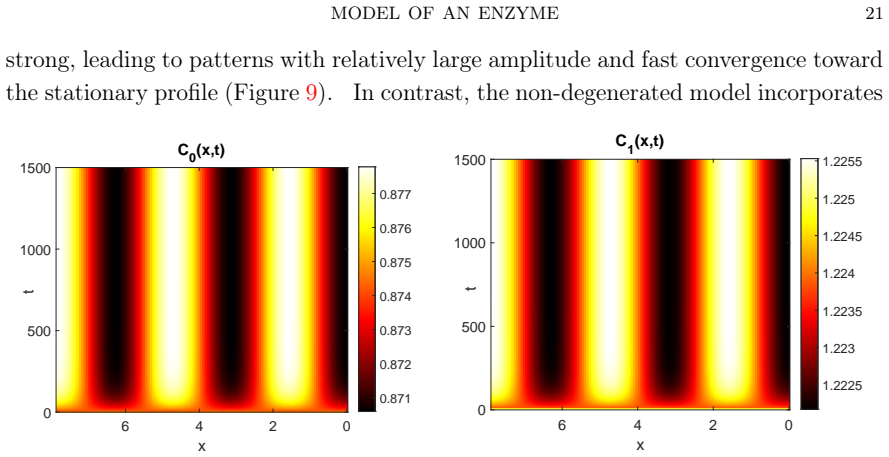


read the original abstract
Spatial organization in metabolic pathways can arise from the interplay between enzymatic reaction kinetics and diffusion-driven instabilities. In this work we investigate how reversible enzyme--substrate binding influences pattern formation in a two-step metabolic pathway. Starting from a mechanistic description in which the substrate reversibly binds to the first enzyme before catalytic conversion, we formulate a three-species reaction--diffusion system that explicitly incorporates the enzyme--substrate complex. We first analyse the homogeneous dynamics and determine the unique steady state of the kinetic system. Exploiting the separation of time scales between the rapid binding kinetics and the slower evolution of metabolite concentrations, we derive a reduced two-variable model using a quasi-steady-state approximation for the enzyme-substrate complex. This reduction preserves the essential nonlinear coupling between catalytic reactions and spatial transport. Linear stability and weakly nonlinear analysis reveal conditions for diffusion-driven (Turing) instability and show that reversible enzyme binding significantly modifies the location and extent of the instability region compared to models with effective kinetics. Numerical simulations confirm the analytical predictions and demonstrate how enzyme-substrate interactions reshape pattern selection and slow the emergence of spatial heterogeneity. These results provide a mechanistic link between enzyme binding kinetics, diffusion-driven pattern formation, and mesoscale metabolic organization. The proposed framework offers a tractable approach for studying spatial patterning in enzymatic networks and may help explain the emergence of structured biochemical domains such as those associated with liquid--liquid phase separation.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript develops a three-species reaction-diffusion model for a two-step metabolic pathway that explicitly includes reversible enzyme-substrate complex formation. After identifying the unique homogeneous steady state, the authors invoke a quasi-steady-state approximation (QSSA) on the complex to obtain a reduced two-variable system. Linear stability analysis together with weakly nonlinear analysis then identifies the conditions for diffusion-driven (Turing) instability and demonstrates that reversible binding shifts the location and extent of the instability region relative to models that employ effective kinetics. Numerical simulations are used to corroborate the analytical predictions and to illustrate effects on pattern selection and the time scale of heterogeneity emergence.
Significance. If the central results are robust, the work supplies a concrete mechanistic link between enzyme-binding kinetics and the parameter regimes that permit spatial patterning in metabolic pathways. The explicit retention of binding steps, rather than immediate reduction to effective rates, is a clear strength; the combination of linear stability, weakly nonlinear analysis, and direct numerical confirmation provides a tractable analytical route that could be applied to larger enzymatic networks. The findings offer a plausible explanation for mesoscale metabolic organization without invoking additional regulatory mechanisms.
major comments (2)
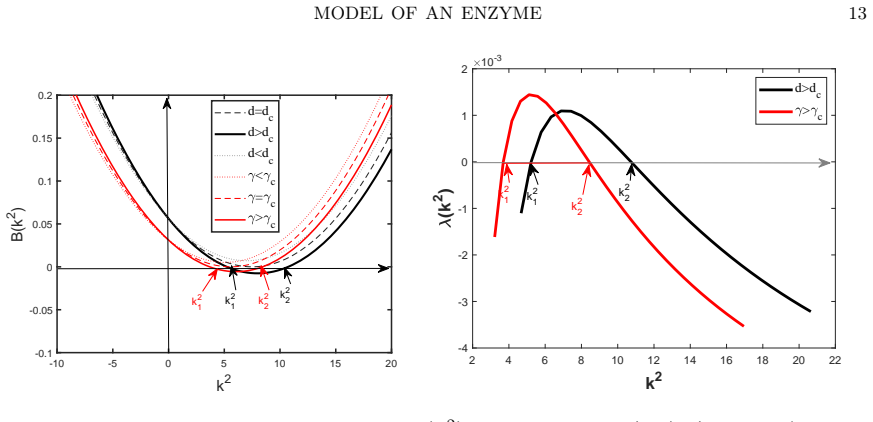
- [§3] §3 (QSSA reduction): The reduction assumes that binding equilibrates much faster than diffusion and metabolite evolution, allowing the complex to be slaved locally. However, the manuscript provides no explicit comparison of the dispersion relation or the critical diffusion ratio between the full three-species system and the reduced two-variable model. Without this comparison or an error bound on the approximation, it remains unclear whether the reported shifts in the instability region are preserved when diffusion of the complex is retained.
- [§4] §4 (linear stability analysis): The claim that reversible enzyme binding 'significantly modifies' the location and extent of the Turing region is central, yet the text does not quantify the magnitude of the shift (e.g., change in critical wave number or minimal diffusion ratio) for representative parameter values. A direct overlay of the instability boundaries for the reduced model versus the effective-kinetics model would make the modification concrete and testable.
minor comments (2)
- Notation for the binding and unbinding rates is introduced without a consolidated table of symbols; adding such a table would improve readability when comparing the full and reduced systems.
- The weakly nonlinear analysis section would benefit from an explicit statement of the amplitude equation coefficients and the conditions under which they remain positive, to allow readers to reproduce the pattern-selection conclusions.
Simulated Author's Rebuttal
We thank the referee for their thorough review and constructive feedback on our manuscript. We appreciate the positive assessment of the significance of our work and address the major comments point by point below.
read point-by-point responses
-
Referee: [§3] §3 (QSSA reduction): The reduction assumes that binding equilibrates much faster than diffusion and metabolite evolution, allowing the complex to be slaved locally. However, the manuscript provides no explicit comparison of the dispersion relation or the critical diffusion ratio between the full three-species system and the reduced two-variable model. Without this comparison or an error bound on the approximation, it remains unclear whether the reported shifts in the instability region are preserved when diffusion of the complex is retained.
Authors: We agree with the referee that an explicit comparison would better justify the QSSA. In the revised version, we will add a section comparing the dispersion relations of the full three-species system and the reduced two-variable model for parameters where the binding time scale is sufficiently separated from diffusion. We will also derive and present error bounds on the approximation to confirm that the reported shifts in the Turing instability region remain valid. revision: yes
-
Referee: [§4] §4 (linear stability analysis): The claim that reversible enzyme binding 'significantly modifies' the location and extent of the Turing region is central, yet the text does not quantify the magnitude of the shift (e.g., change in critical wave number or minimal diffusion ratio) for representative parameter values. A direct overlay of the instability boundaries for the reduced model versus the effective-kinetics model would make the modification concrete and testable.
Authors: We acknowledge that providing quantitative measures and a visual comparison would strengthen the central claim. We will revise the manuscript to include an overlay plot of the instability boundaries in the (diffusion ratio, wave number) plane for both the reduced model with explicit binding and the effective-kinetics model. Additionally, we will report specific numerical values for the shifts in critical parameters, such as the minimal diffusion ratio required for instability, using representative parameter sets. revision: yes
Circularity Check
Derivation chain is self-contained; no reductions to inputs by construction
full rationale
The paper begins with an explicit three-species mechanistic reaction-diffusion system incorporating reversible enzyme-substrate binding. It invokes the standard quasi-steady-state approximation justified by explicit time-scale separation between fast binding and slow metabolite evolution, yielding a reduced two-variable model whose Jacobian and dispersion relation are then analyzed for Turing instability. The reported modification of the instability region relative to effective-kinetics models is obtained by direct comparison of these derived dispersion relations; it is not a fitted parameter renamed as a prediction, nor does any step reduce to a self-definition or self-citation chain. The derivation remains independent of the target result and is externally falsifiable via the full three-species system or numerical simulation.
Axiom & Free-Parameter Ledger
free parameters (2)
- binding and unbinding rate constants
- diffusion coefficients
axioms (2)
- domain assumption Quasi-steady-state approximation holds for the enzyme-substrate complex
- standard math Mass-action kinetics govern the reactions
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
We derive a reduced two-variable model using a quasi-steady-state approximation for the enzyme-substrate complex... Linear stability and weakly nonlinear analysis reveal conditions for diffusion-driven (Turing) instability
-
IndisputableMonolith/Foundation/AlexanderDuality.leanalexander_duality_circle_linking unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
the non-degenerated system (3.3) has the same structure as the primary model (1.1), but with modified effective coefficients
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Reference graph
Works this paper leans on
-
[1]
Bending the rules: curvature’s impact on cell biology.BMC biology, 23(1):296, 2025
Carole Arnold, Ismail Tahmaz, Marie-Ly Chapon, Hasna Maayouf, Valeriy Luchnikov, Jean-Louis Milan, Fabricio Borghi, and Laurent Pieuchot. Bending the rules: curvature’s impact on cell biology.BMC biology, 23(1):296, 2025. 4 MODEL OF AN ENZYME 25
work page 2025
-
[2]
Salman F Banani, Hyun O Lee, Anthony A Hyman, and Michael K Rosen. Biomolecular condensates: organizers of cellular biochemistry.Nature reviews Molecular cell biology, 18(5):285–298, 2017. 4
work page 2017
-
[3]
Alessandro Bevilacqua, Mauricio Rios Maciel, Mark V Sullivan, Stefano Pascarelli, Mirco Dindo, Amy Q Shen, and Paola Laurino. Enzyme activity regulates sub- stratediffusionbymodulatingviscosityincrowdedmilieu.BioRxiv, pages2024–09,
-
[4]
Alexander Buchner, Filipe Tostevin, and Ulrich Gerland. Clustering and optimal arrangement of enzymes in reaction-diffusion systems.Physical Review Letters, 110(20):208104, 2013. 2
work page 2013
-
[5]
Michele Castellana, Maxwell Z Wilson, Yifan Xu, Preeti Joshi, Ileana M Cristea, Joshua D Rabinowitz, Zemer Gitai, and Ned S Wingreen. Enzyme clustering accel- erates processing of intermediates through metabolic channeling.Nature biotech- nology, 32(10):1011–1018, 2014. 2, 4, 5
work page 2014
-
[6]
Huiyi Chen, Shunyi Huang, Longcheng Quan, Caiyuan Yu, Yang Zhu, Xiaocong Sun, Yuanli Zhang, Liehua Deng, and Feng Chen. Liquid–liquid phase separation: a potentially fundamental mechanism of sepsis.Cell Death Discovery, 11(1):310,
-
[7]
Zhihao Chen, Ying Huai, Wenjing Mao, Xuehao Wang, Kang Ru, Airong Qian, and Hong Yang. Liquid–liquid phase separation of biomacromolecules and its roles in metabolic diseases.Cells, 11(19):3023, 2022. 2
work page 2022
-
[8]
PhD thesis, Johns Hopkins University, 2022
Basilio Cieza Huaman.Investigation of Molecular condensates using a Reaction- Diffusion Master Equation Model. PhD thesis, Johns Hopkins University, 2022. 2
work page 2022
-
[9]
Chemical oscillators in structured media
Irving R Epstein, Vladimir K Vanag, Anna C Balazs, Olga Kuksenok, Pratyush Dayal, and Amitabh Bhattacharya. Chemical oscillators in structured media. Accounts of chemical research, 45(12):2160–2168, 2012. 2
work page 2012
-
[10]
Chikara Furusawa and Kunihiko Kaneko. Emergence of multicellular organisms with dynamic differentiation and spatial pattern.Artificial life, 4(1):79–93, 1998. 2
work page 1998
-
[11]
Effects of cross-diffusion on Turing patterns in a reaction-diffusion Schnakenberg model
G Gambino, S Lupo, and M Sammartino. Effects of cross-diffusion on turing pat- ternsinareaction-diffusionschnakenbergmodel.arXiv preprint arXiv:1501.04890,
work page internal anchor Pith review Pith/arXiv arXiv
-
[12]
Gaetana Gambino, Maria Carmela Lombardo, and Marco Sammartino. Turing instability and traveling fronts for a nonlinear reaction–diffusion system with cross- diffusion.Mathematics and Computers in Simulation, 82(6):1112–1132, 2012. 18
work page 2012
-
[13]
Gaetana Gambino, Maria Carmela Lombardo, and Marco Sammartino. Pattern 26 F. F ARIV AR formation driven by cross-diffusion in a 2d domain.Nonlinear Analysis: Real World Applications, 14(3):1755–1779, 2013. 6
work page 2013
-
[14]
Pattern selection in the 2d fitzhugh–nagumo model.Ricerche di Matem- atica, 68(2):535–549, 2019
Gaetana Gambino, Maria Cristina Lombardo, Gino Rubino, and Marcello Sam- martino. Pattern selection in the 2d fitzhugh–nagumo model.Ricerche di Matem- atica, 68(2):535–549, 2019. 6
work page 2019
-
[15]
Valeria Giunta, Maria Carmela Lombardo, and Marco Sammartino. Pattern for- mationandtransitiontochaosinachemotaxismodelofacuteinflammation.SIAM Journal on Applied Dynamical Systems, 20(4):1844–1881, 2021. 2
work page 2021
-
[16]
Jasmin Hafner and Vassily Hatzimanikatis. Nicepath: Finding metabolic pathways in large networks through atom-conserving substrate–product pairs.Bioinformat- ics, 37(20):3560–3568, 2021. 2
work page 2021
-
[17]
Camilla Beate Hill, Tobias Czauderna, Matthias Klapperstück, Ute Roessner, and Falk Schreiber. Metabolomics, standards, and metabolic modeling for synthetic biology in plants.Frontiers in Bioengineering and Biotechnology, 3:167, 2015. 2
work page 2015
-
[18]
A user’s guide to pde models for chemotaxis
Thomas Hillen and Kevin J Painter. A user’s guide to pde models for chemotaxis. Journal of mathematical biology, 58(1):183–217, 2009. 4
work page 2009
-
[19]
Anthony A Hyman, Christoph A Weber, and Frank Jülicher. Liquid-liquid phase separation in biology.Annual review of cell and developmental biology, 30(1): 39–58, 2014. 4
work page 2014
-
[20]
James Keener and James Sneyd.Mathematical physiology: II: Systems physiology. Springer, 2009. 5, 6
work page 2009
-
[21]
Prentice hall Upper Saddle River, NJ, 2002
Hassan K Khalil and Jessy W Grizzle.Nonlinear systems, volume 3. Prentice hall Upper Saddle River, NJ, 2002
work page 2002
-
[22]
Jinyoung Kim, Sean D Lawley, and Jinsu Kim. A reaction network model of microscale liquid–liquid phase separation reveals effects of spatial dimension.The Journal of Chemical Physics, 161(20), 2024. 2
work page 2024
-
[23]
Georg Krainer, Timothy J Welsh, Jerelle A Joseph, Peter St George-Hyslop, An- thony A Hyman, Rosana Collepardo-Guevara, Simon Alberti, and Tuomas PJ Knowles. Reentrant liquid condensate phase of proteins is stabilized by hydropho- bic and non-ionic interactions.Biophysical Journal, 120(3):28a, 2021. 2
work page 2021
-
[24]
Hyo Lee, Saet Buyl Lee, Sangkyu Park, Jaeeun Song, and Beom-Gi Kim. Biochem- ical evaluation of molecular parts for flavonoid production using plant synthetic biology.Frontiers in Plant Science, 16:1528122, 2025. 2
work page 2025
-
[25]
Guangle Li, Chengqian Yuan, and Xuehai Yan. Peptide-mediated liquid–liquid phase separation and biomolecular condensates.Soft Matter, 21(10):1781–1812,
-
[26]
Samuel Lim and Douglas S Clark. Phase-separated biomolecular condensates for MODEL OF AN ENZYME 27 biocatalysis.Trends in Biotechnology, 42(4):496–509, 2024. 2
work page 2024
-
[27]
Zheran Liu, Zijian Qin, Yingtong Liu, Xi Xia, Ling He, Na Chen, Xiaolin Hu, and Xingchen Peng. Liquid–liquid phase separation: Roles and implications in future cancer treatment.International Journal of Biological Sciences, 19(13):4139, 2023. 2
work page 2023
-
[28]
Jiahua Lu, Junjie Qian, Zhentian Xu, Shengyong Yin, Lin Zhou, Shusen Zheng, and Wu Zhang. Emerging roles of liquid–liquid phase separation in cancer: from protein aggregation to immune-associated signaling.Frontiers in cell and devel- opmental biology, 9:631486, 2021. 2
work page 2021
-
[29]
The turing model for biological pattern formation
Philip K Maini and Thomas E Woolley. The turing model for biological pattern formation. InThe dynamics of biological systems, pages 189–204. Springer, 2019. 2
work page 2019
-
[30]
Philip K Maini, Thomas E Woolley, Ruth E Baker, Eamonn A Gaffney, and S Seirin Lee. Turing’s model for biological pattern formation and the robustness problem.Interface focus, 2(4):487–496, 2012. 2
work page 2012
-
[31]
Katarina Milicevic, Branislava Rankovic, Pavle R Andjus, Danijela Bataveljic, and Dragomir Milovanovic. Emerging roles for phase separation of rna-binding proteins in cellular pathology of als.Frontiers in cell and developmental biology, 10:840256, 2022. 2
work page 2022
-
[32]
A reaction-diffusion model captures the essence of liquid-liquid phase separation
Nayana Mukherjee, Abdul Wasim, Jagannath Mondal, and Pushpita Ghosh. A reaction-diffusion model captures the essence of liquid-liquid phase separation. bioRxiv, pages 2024–04, 2024. 2
work page 2024
-
[33]
James D Murray.Mathematical biology: I. An introduction, volume 17. Springer Science & Business Media, 2007. 6
work page 2007
-
[34]
Yusuke Nakasone and Masahide Terazima. A time-resolved diffusion technique for detection of the conformational changes and molecular assembly/disassembly processes of biomolecules.Frontiers in Genetics, 12:691010, 2021. 2
work page 2021
-
[35]
PhD thesis, Politecnico di Torino, 2022
Gabriele Rigano.Models of cell membrane patterning via diffusion-controlled phase separation. PhD thesis, Politecnico di Torino, 2022. 2
work page 2022
-
[36]
Barbara Schamberger, Ricardo Ziege, Karine Anselme, Martine Ben Amar, Michał Bykowski, André PG Castro, Amaia Cipitria, Rhoslyn A Coles, Rumiana Dimova, Michaela Eder, et al. Curvature in biological systems: its quantification, emer- gence, and implications across the scales.advanced materials, 35(13):2206110,
-
[37]
A. M. Turing. The chemical basis of morphogensis.Phil. Trans. Roy. Soc., B (237):37–72, 1952. 2
work page 1952
-
[38]
David Zwicker, Rabea Seyboldt, Christoph A Weber, Anthony A Hyman, and 28 F. F ARIV AR Frank Jülicher. Growth and division of active droplets provides a model for pro- tocells.Nature Physics, 13(4):408–413, 2017. 4, 5 1 Institute of Atmospheric Sciences and Climate (CNR-ISAC), Rome, Italy Email address:faezehfarivar@cnr.it
work page 2017
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.