Recognition: unknown
Building and maintaining a System of Intracellular Compartments
Pith reviewed 2026-05-10 13:49 UTC · model grok-4.3
The pith
Vesicular transport and cisternal progression are two dynamical phases of one nonequilibrium fusion-fission process.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
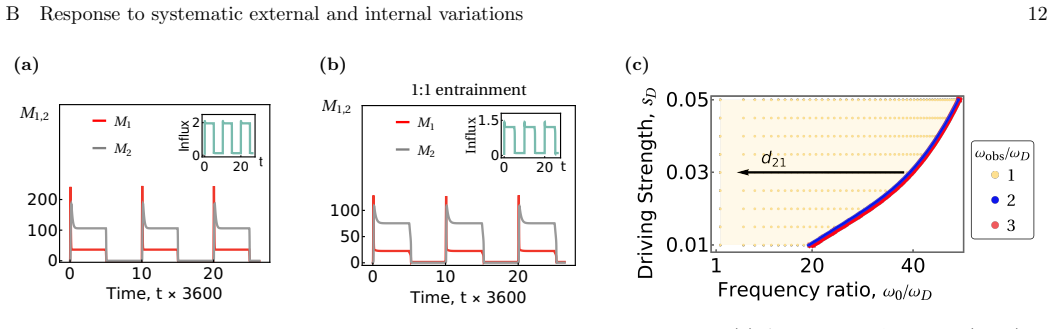
Organelle patterning and heritability arise from nonequilibrium mechanochemical fusion-fission cycles that violate detailed balance. Within this dynamical-systems description the Golgi and endosomes exhibit distinct regimes, including fixed-point stability, cell-cycle-timed limit cycles, and cisternal progression; the two classical models of Golgi organization are therefore alternative phases of one continuous stochastic process rather than competing mechanisms.
What carries the argument
A stochastic mechanochemical fusion-fission model that violates detailed balance, whose dynamical regimes (fixed points, limit cycles, progressing states) map onto biological phenotypes of stable cisternae, periodic reassembly, and cisternal maturation.
If this is right
- Stable cisternae correspond to a fixed-point regime of the fusion-fission dynamics.
- Periodic cell-cycle-dependent dissolution and reassembly corresponds to a limit-cycle regime with definite phase relations.
- Cisternal progression emerges as a separate dynamical phase of the same nonequilibrium process.
- Modulating the interplay between glycosylation enzymes and fission-fusion rates can control both the chemical identity and the number of cisternae.
- The system exhibits definite, testable responses to systematic perturbations or driving protocols.
Where Pith is reading between the lines
- Cells may achieve rapid switching between transport modes by tuning only a few global rate parameters rather than by deploying separate molecular machineries.
- Similar nonequilibrium fusion-fission frameworks could unify patterning mechanisms in other membrane-bound organelles.
- The model suggests that mutations or drugs altering fission-fusion balance should produce predictable transitions between Golgi phenotypes that can be checked in live-cell imaging.
Load-bearing premise
The complex intracellular membrane dynamics can be captured by a stochastic fusion-fission model that violates detailed balance and whose regimes map directly to observed phenotypes without extra molecular details.
What would settle it
An experiment that systematically varies the ratio of fusion to fission rates or glycosylation enzyme activity and records whether the Golgi switches between stable cisternae and progressive maturation exactly as the phase diagram of the model predicts.
Figures






read the original abstract
Organelle patterning and its heritability remain central mysteries in cell biology, highlighting the fundamental tension between genetic inheritance and self-assembly. Here, we explore the nonequilibrium assembly and size control of the Golgi complex and endosomes, amid a continuous flux of membrane traffic, within a stochastic framework of mechanochemical fusion-fission cycles that violate detailed balance. Using a dynamical systems approach, we identify distinct, robust regimes, ranging from fixed points to limit cycles with definite phase relations. We identify these dynamical regimes with diverse phenotypes, from stable cisternae to periodic, cell-cycle-dependent dissolution/reassembly to cisternal progression. We analyse its dynamic response to systematic perturbations or driving protocols and make definite predictions that may be tested experimentally. Our analysis reveals that the two competing models of Golgi organization-vesicular transport and cisternal progression - are, in fact, two phases of the same underlying nonequilibrium process. Finally, our framework offers a strategy for controlling cisternal chemical identity and number and by modulating the interplay between glycosylation enzymes and membrane fission-fusion dynamics.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript develops a stochastic mechanochemical model of fusion-fission cycles that violate detailed balance to describe the nonequilibrium assembly, size control, and maintenance of the Golgi complex and endosomes under continuous membrane traffic. Using a dynamical systems approach, it identifies robust regimes ranging from fixed points to limit cycles with definite phase relations, mapping these to biological phenotypes including stable cisternae, cell-cycle-dependent dissolution/reassembly, and cisternal progression. The central claim is that the competing models of vesicular transport and cisternal progression are two phases of the same underlying process, with additional analysis of responses to perturbations, definite experimental predictions, and a strategy for controlling cisternal chemical identity via enzyme-membrane coupling.
Significance. If the claimed direct mapping from coarse-grained fusion-fission dynamics to the two Golgi organization models holds without requiring extra molecular rules, the work would unify two longstanding competing frameworks and provide a nonequilibrium self-assembly perspective on organelle patterning and heritability. The identification of limit cycles and phase relations, together with perturbation protocols and falsifiable predictions, represents a strength; however, the significance is limited by the absence of explicit parameter values, simulation details, or quantitative data comparisons in the presented framework.
major comments (2)
- [Results on dynamical regimes and phase diagram] The central unification claim (that vesicular transport and cisternal progression are distinct phases of the same stochastic process) is load-bearing but rests on the assumption that the two-parameter mechanochemical model (fusion/fission rates and enzyme-membrane coupling strengths) alone suffices to reproduce essential flux patterns and chemical-identity maintenance. This requires explicit demonstration that no additional rules (e.g., coat-protein specificity or SNARE-mediated targeting) are needed for the regimes to map uniquely to the biological phenotypes; otherwise the equivalence reduces to model construction rather than derivation.
- [Dynamical systems analysis and phenotype identification] The mapping of fixed points to stable cisternae/vesicular transport and limit cycles to cisternal progression is presented without quantitative anchoring to biological observables (e.g., measured fusion rates, cisternal lifetimes, or glycosylation kinetics). Without such anchoring or comparison to existing data, it is unclear whether the identified regimes emerge independently or are selected by parameter choices that encode the target phenotypes.
minor comments (2)
- [Abstract and Introduction] The abstract and introduction would benefit from a brief statement of the exact stochastic update rules and the form of the detailed-balance violation to allow readers to assess the minimal assumptions immediately.
- [Model definition] Notation for the mechanochemical coupling terms and the definition of the phase relations in the limit-cycle regime should be clarified with an explicit equation or diagram early in the methods or results.
Simulated Author's Rebuttal
We thank the referee for their insightful comments on our manuscript. We address each major comment below and have made revisions to clarify and strengthen the presentation of our results.
read point-by-point responses
-
Referee: [Results on dynamical regimes and phase diagram] The central unification claim (that vesicular transport and cisternal progression are distinct phases of the same stochastic process) is load-bearing but rests on the assumption that the two-parameter mechanochemical model (fusion/fission rates and enzyme-membrane coupling strengths) alone suffices to reproduce essential flux patterns and chemical-identity maintenance. This requires explicit demonstration that no additional rules (e.g., coat-protein specificity or SNARE-mediated targeting) are needed for the regimes to map uniquely to the biological phenotypes; otherwise the equivalence reduces to model construction rather than derivation.
Authors: Our model is constructed as a minimal description of fusion-fission cycles with mechanochemical coupling, deliberately omitting molecular details like coat proteins and SNAREs to test whether the core process can generate the observed behaviors. We demonstrate that within this framework, the fixed-point and limit-cycle regimes emerge robustly and map to the vesicular transport and cisternal progression phenotypes, respectively. This shows that additional rules are not required for the qualitative unification. In the revised manuscript, we have expanded the discussion to explicitly state that molecular specificities would enter as effective parameters in our rates, and we provide examples of how the model remains consistent with known biology without them. We have also added a supplementary figure illustrating the robustness of the phase diagram to small perturbations that might mimic such rules. revision: yes
-
Referee: [Dynamical systems analysis and phenotype identification] The mapping of fixed points to stable cisternae/vesicular transport and limit cycles to cisternal progression is presented without quantitative anchoring to biological observables (e.g., measured fusion rates, cisternal lifetimes, or glycosylation kinetics). Without such anchoring or comparison to existing data, it is unclear whether the identified regimes emerge independently or are selected by parameter choices that encode the target phenotypes.
Authors: We acknowledge that our original presentation focused on the qualitative dynamical regimes. To address this, we have revised the manuscript to include explicit estimates of parameter values drawn from the literature, such as typical fusion rates in the range of 0.1-1 events per minute per cisterna and cisternal lifetimes of 10-30 minutes. We show that the limit cycle regime corresponds to these values, with phase relations matching observed progression times. A new table compares model predictions to experimental observables, and we discuss how the regimes arise from the bifurcation structure rather than being tuned to specific phenotypes. While a full quantitative fit to all data is beyond the scope of this theoretical work, these additions provide the requested anchoring. revision: yes
Circularity Check
No significant circularity; derivation self-contained
full rationale
The abstract describes a stochastic mechanochemical fusion-fission model violating detailed balance, followed by dynamical-systems identification of regimes (fixed points to limit cycles) that are then associated with phenotypes including vesicular transport and cisternal progression. This mapping is presented as an outcome of the analysis rather than a definitional equivalence or a fitted parameter renamed as prediction. No equations, self-citations, or ansatzes are quoted that would reduce the central unification claim to the model's inputs by construction. The framework is offered as making testable predictions, indicating the regimes are derived independently of the target biological interpretations.
Axiom & Free-Parameter Ledger
free parameters (2)
- fusion and fission rates
- enzyme-membrane coupling strengths
axioms (2)
- domain assumption Mechanochemical fusion-fission cycles violate detailed balance
- domain assumption Continuous membrane traffic flux can be modeled as stochastic cycles
Reference graph
Works this paper leans on
-
[1]
Solve the differential equations Eqs.(S68),(S69) in(𝑀 1, 𝑀2)(Eqs.(12),(13) in the main text) for given values of parameters and initial conditions
-
[2]
We compute the following quantities :|𝑀 1 (𝜏𝑎) − 𝑀1 (𝜏𝑐)|,|𝑀 1 (𝜏𝑏) −𝑀 1 (𝜏𝑑)|,|𝑀 2 (𝜏𝑎) −𝑀 2 (𝜏𝑐)|,|𝑀 2 (𝜏𝑐) −𝑀 2 (𝜏𝑑)|, 𝑀 1 (𝑡=𝜏 𝑒), 𝑀2 (𝑡=𝜏 𝑒)
Choose, say, five time points𝜏 𝑎 < 𝜏 𝑏 < 𝜏 𝑐 < 𝜏 𝑑 < 𝜏 𝑒, these time points are taken to be large enough numbers to make sure that the system has arrived at a steady-state. We compute the following quantities :|𝑀 1 (𝜏𝑎) − 𝑀1 (𝜏𝑐)|,|𝑀 1 (𝜏𝑏) −𝑀 1 (𝜏𝑑)|,|𝑀 2 (𝜏𝑎) −𝑀 2 (𝜏𝑐)|,|𝑀 2 (𝜏𝑐) −𝑀 2 (𝜏𝑑)|, 𝑀 1 (𝑡=𝜏 𝑒), 𝑀2 (𝑡=𝜏 𝑒)
-
[3]
The steady-state is a fixed point solution if 0<|𝑀 1 (𝜏𝑎) −𝑀 1 (𝜏𝑐)|< 𝛿 1, 0<|𝑀 1 (𝜏𝑏) −𝑀 1 (𝜏𝑑)|< 𝛿 1, 0<|𝑀 2 (𝜏𝑎) − 𝑀2 (𝜏𝑐)|< 𝛿 1, 0<|𝑀 2 (𝜏𝑏) −𝑀 2 (𝜏𝑑)|< 𝛿 1,𝑀 1 (𝑡=𝜏 𝑒)< 𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ and𝑀 2 (𝑡=𝜏 𝑒)< 𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ ,𝛿 1 is the tolerance assumed and𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ =300 is the threshold size assumed for the stable state
-
[4]
The steady-state is a limit cycle solution if𝛿 1 ≤ |𝑀 1 (𝜏𝑎) −𝑀 1 (𝜏𝑐)|< 𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ ,𝛿 1 ≤ |𝑀 1 (𝜏𝑏) −𝑀 1 (𝜏𝑑)|< 𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ , 𝛿1 ≤ |𝑀 2 (𝜏𝑎) −𝑀 2 (𝜏𝑐)|< 𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ ,𝛿 1 ≤ |𝑀 2 (𝜏𝑏) −𝑀 2 (𝜏𝑑)|< 𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ ,𝑀 1 (𝑡=𝜏 𝑒)< 𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ and𝑀 2 (𝑡=𝜏 𝑒)< 𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ
-
[5]
S5.5 Arriving at the phase diagram and phase boundaries for the two-cisternae system 47 No
The system is unbounded if𝑀 1 (𝑡=𝜏 𝑒) ≥𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ or𝑀 2 (𝑡=𝜏 𝑒) ≥𝑀 ℎ𝑖𝑔ℎ 𝑡 ℎ . S5.5 Arriving at the phase diagram and phase boundaries for the two-cisternae system 47 No. of distinct roots Algebraic capacity 0 1 2 3 4 5 0 10 20 30 Influx rate, v Root count, r N rN =18 Figure S14:Structural stability of two-cisternae dynamical system. The structural stabilit...
-
[6]
mean behaviour
While the numerical scheme presented above is adequate for the present purposes, it is subject to the following limitations – (i) it would not detect limit cycle solutions with a radius𝑟 𝜏, such that𝑟 𝜏 < 𝛿 1 or𝑟 𝜏 > 𝑀 𝑡 ℎ – and would assume them to be fixed point solutions. (ii) Fixed point solutions that reach the asymptotic value at a time much larger ...
-
[7]
We have usedMathematicafor this (which uses Buchberger algorithm and DegreeReverseLexicographic order, see [86] for details)
Compute the Gr¨ obner basis from the system of equations/vector fields (e.g.𝑥 3 +𝑏 𝑥+𝑐for cusp singularity [47], where x is the state variable and b and c are constant parameters). We have usedMathematicafor this (which uses Buchberger algorithm and DegreeReverseLexicographic order, see [86] for details)
-
[8]
Extract the leading monomials from each polynomial in the Gr¨ obner basis (e.g.,{𝑥 3})
-
[9]
Generate all monomials up to a chosen maximum polynomial Degree:{1, 𝑥, 𝑥 2, 𝑥3, 𝑥4, 𝑥5, . . .}
-
[10]
Reject all monomials divisible by any leading monomial (e.g., reject𝑥 3, 𝑥4, 𝑥5, . . .)
-
[11]
The surviving monomials form the standard monomial basis (e.g.,{1, 𝑥, 𝑥 2})
-
[12]
Algebraic capacity = dimension of standard monomial basis (e.g., capacity = 3)
-
[13]
Choose specific numerical values for the parameters and numerically solve the system to count the number of distinct complex roots (geometric roots)
-
[14]
Compare: if geometric roots = algebraic capacity, the system is structurally stable. 48
-
[15]
The above steps for the 2-cisternae system are tabulated in Table S4
If geometric roots<algebraic capacity, the system is structurally unstable. The above steps for the 2-cisternae system are tabulated in Table S4. The number of distinct complex roots and the algebraic capacity are plotted in Fig. S14. Vector field ( 𝑣 𝑎1 + 𝑀2 1 𝐶11 +𝑀 2 1 ! − 𝑑1 𝑀1 𝐶12 +𝑀 1 − 𝑑12 𝑀1 𝐶12 +𝑀 1 𝑎2 + 𝑀2 2 𝐶21 +𝑀 2 2 ! + 𝑑21 𝑀2 𝐶22 +𝑀 2 𝑎1 + 𝑀...
2000
-
[16]
Parameter values,𝑣=2, 𝐶11 =100,𝐶 22 =20,𝐶 12 =20,𝑑 12 =2,𝑑 21 =0,𝑑 1 =1,𝑑 2 =1.5,𝑎 1 =𝑎 2 =0.05
+ ∑︁ 𝑖 𝜏𝑖 𝑆 𝑁 (𝐸 1 𝑖 , 𝐸2 𝑖 , 𝑥1 𝑖 , 𝑥2 𝑖 ) 𝑖∈SN bifurcations encompassed by the trajectory (S105) S8.2 De novo assembly of two cisternae 55 (a) M1 M2 0 500 1000 1500 0 50 100 150 200 Time, t M1,2 τSN τSN+τf (b) (c) M1 M2 0 500 1000 15000 20 40 Time, t M2 τSN τSN+τ0 (d) Figure S18:Formation time for two cisternae.(a) Cisternae formation time for fixed poi...
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.