Recognition: unknown
Embedded DNA Inference in In-Body Nanonetworks: Detection, Delay, and Communication Trade-Offs
Pith reviewed 2026-05-10 13:16 UTC · model grok-4.3
The pith
Simulations show embedded DNA inference at nanonodes improves detection of weak-to-moderate anomalies while keeping communication costs competitive.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
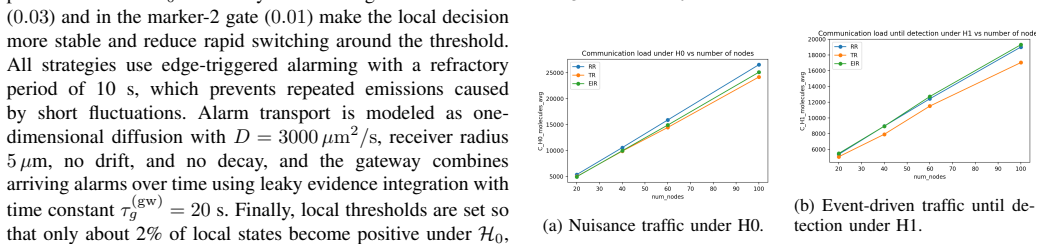
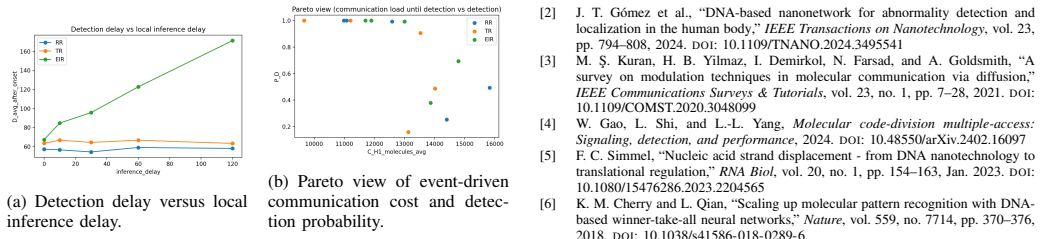
The simulations identify a bounded EIR success regime in the weak-to-moderate anomaly range: EIR can improve detection relative to RR and TR while remaining competitive in event-driven communication cost, especially relative to RR. The gain does not come from uniformly lower activity, but from more stable local alarm dynamics. EIR does not dominate globally; TR often remains cheaper when abnormalities are present, and EIR incurs additional local delay.
What carries the argument
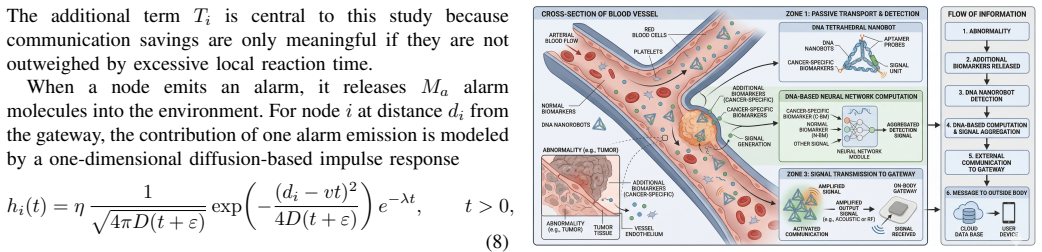
Embedded inference reporting (EIR) using DNA strand-displacement-based computation with marker gating, edge-triggered alarming, hysteretic state transitions, temporally correlated marker dynamics, diffusion-based alarm transport, and leaky gateway evidence integration.
If this is right
- EIR improves detection performance over raw reporting and threshold reporting specifically in weak-to-moderate anomaly conditions.
- Communication costs with EIR stay competitive with raw reporting because of more stable local alarm dynamics rather than reduced overall activity.
- Threshold reporting often incurs lower costs than EIR once strong abnormalities are present.
- EIR adds measurable local processing delay that must be weighed against its detection improvements in the target regime.
Where Pith is reading between the lines
- The limited success regime suggests hybrid schemes that switch reporting methods according to anomaly strength could optimize overall performance.
- The emphasis on local stability points to potential use in other molecular communication settings where diffusion noise makes direct transmission unreliable.
- Testing the DNA abstraction in controlled microfluidic experiments with actual strand-displacement gates would clarify how far the simulated bounds apply in practice.
Load-bearing premise
The communication-oriented abstraction of DNA strand-displacement computation and diffusion transport accurately captures real biological conditions and dynamics.
What would settle it
A wet-lab experiment showing that actual DNA strand-displacement circuits in a diffusive medium fail to produce the stable hysteretic alarm states predicted for moderate marker correlations would disprove the reported detection gains.
Figures

read the original abstract
In-body molecular nanonetworks promise early abnormality detection close to the source of biochemical events, but their communication capabilities are severely constrained by slow diffusion-based signaling and unstable alarm traffic. We study whether simple embedded DNA-based inference at the nanonode can improve alarm transmission to an external gateway. We compare raw reporting (RR), single-marker threshold reporting (TR), and embedded inference reporting (EIR) under a communication-oriented abstraction of DNA strand-displacement-based computation with marker gating, edge-triggered alarming, hysteretic state transitions, temporally correlated marker dynamics, diffusion-based alarm transport, and leaky gateway evidence integration. The simulations identify a bounded EIR success regime in the weak-to-moderate anomaly range: EIR can improve detection relative to RR and TR while remaining competitive in event-driven communication cost, especially relative to RR. The gain does not come from uniformly lower activity, but from more stable local alarm dynamics. EIR does not dominate globally; TR often remains cheaper when abnormalities are present, and EIR incurs additional local delay. These results point to a limited operating regime in which EIR is useful, rather than to a general advantage across settings.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper claims that embedded DNA inference reporting (EIR) using a communication-oriented abstraction of strand-displacement computation (marker gating, edge-triggered hysteretic transitions, temporally correlated markers, diffusion transport, leaky integration) improves abnormality detection over raw reporting (RR) and threshold reporting (TR) in a bounded weak-to-moderate anomaly regime, while remaining competitive on event-driven communication cost due to more stable local alarm dynamics rather than uniformly lower activity; EIR does not dominate globally and adds local delay.
Significance. If the simulation results hold, the work would be significant for molecular nanonetwork design by identifying a limited operating regime where simple embedded inference provides detection gains without general superiority, highlighting trade-offs in detection, delay, and cost for diffusion-based systems aimed at early abnormality detection.
major comments (2)
- [Abstract] Abstract and simulation description: the central claim of a bounded EIR success regime in the weak-to-moderate range rests on Monte Carlo simulations, yet no parameter values, statistical methods, sensitivity analysis, or validation against measured strand-displacement kinetics or in-vivo diffusion coefficients are provided; without these, the location of the regime and the qualitative ordering versus RR/TR cannot be assessed for robustness and may be artifacts of the stylized model.
- [Abstract] The communication-oriented abstraction (marker gating, edge-triggered alarming, hysteretic state transitions, temporally correlated marker dynamics, diffusion-based alarm transport, leaky gateway integration) is load-bearing for all reported trade-offs, but no comparison to real biological noise levels, leak rates, or stochastic reaction-diffusion validation is described; if the abstraction over-smooths noise or underestimates leaks, the reported stability advantage reverses.
Simulated Author's Rebuttal
We thank the referee for the constructive and detailed review, which highlights key aspects of robustness and model fidelity. We address each major comment point by point below, agreeing where revisions are needed to strengthen the presentation of parameters, statistics, and abstraction validation. The revisions will focus on clarifying existing content and adding new analysis without altering the core simulation results.
read point-by-point responses
-
Referee: [Abstract] Abstract and simulation description: the central claim of a bounded EIR success regime in the weak-to-moderate range rests on Monte Carlo simulations, yet no parameter values, statistical methods, sensitivity analysis, or validation against measured strand-displacement kinetics or in-vivo diffusion coefficients are provided; without these, the location of the regime and the qualitative ordering versus RR/TR cannot be assessed for robustness and may be artifacts of the stylized model.
Authors: We thank the referee for this observation. The full manuscript contains a Simulation Setup section (Section IV) that specifies the Monte Carlo configuration (10,000 runs per parameter combination with 95% confidence intervals on detection probability and cost metrics), explicit parameter values (diffusion coefficient D = 5×10^{-10} m²/s, strand-displacement forward rate k = 10^6 M^{-1}s^{-1}, marker correlation time τ = 10 s, gateway leak rate λ = 0.01 s^{-1}, and anomaly strength parameterized as concentration ratios from 1.05 to 4.0), and the statistical procedure for identifying the bounded regime. These details were not foregrounded in the abstract. We will revise the abstract to reference the parameter ranges and add a dedicated sensitivity analysis subsection that perturbs the five most influential parameters by ±30% and confirms the weak-to-moderate regime boundaries remain qualitatively stable. Direct experimental validation against new in-vivo measurements is outside the scope of this modeling study; we will instead add citations to published kinetic and diffusion data that informed the chosen values. revision: yes
-
Referee: [Abstract] The communication-oriented abstraction (marker gating, edge-triggered alarming, hysteretic state transitions, temporally correlated marker dynamics, diffusion-based alarm transport, leaky gateway integration) is load-bearing for all reported trade-offs, but no comparison to real biological noise levels, leak rates, or stochastic reaction-diffusion validation is described; if the abstraction over-smooths noise or underestimates leaks, the reported stability advantage reverses.
Authors: We agree that the abstraction carries the reported trade-offs. The model already encodes Poisson shot noise in marker production, exponential autocorrelation for temporal marker correlation, and an explicit leaky integrator at the gateway (λ = 0.01 s^{-1}). However, we did not include side-by-side numerical comparisons to empirical biological noise spectra or leak rates. In revision we will insert a new table in Section III that maps each abstraction element to representative values from the DNA strand-displacement and molecular-communication literature (leak rates 10^{-4}–10^{-1} s^{-1}, diffusion coefficients 10^{-11}–10^{-9} m²/s in tissue). We will also add a short stochastic reaction-diffusion validation using a simplified Gillespie simulation on a reduced geometry to verify that the continuous abstraction does not materially under-estimate leak-induced false alarms. If measured leak rates prove substantially higher than our nominal value, the stability advantage narrows, and we will state this limitation explicitly in the conclusions. revision: yes
Circularity Check
No circularity: results from direct simulation comparisons
full rationale
The paper reports Monte Carlo simulation outcomes for three explicit strategies (raw reporting, threshold reporting, embedded inference reporting) under a fixed communication-oriented abstraction of strand-displacement dynamics, gating, hysteresis, diffusion, and leaky integration. No equations, fitted parameters, or self-citations are used to derive the reported EIR success regime; the bounded regime and relative performance metrics emerge directly from the simulation runs rather than by algebraic reduction or renaming of inputs. The work is therefore self-contained against external benchmarks and exhibits none of the enumerated circularity patterns.
Axiom & Free-Parameter Ledger
axioms (2)
- domain assumption DNA strand-displacement computation can be abstracted via marker gating, edge-triggered alarming, hysteretic state transitions, and temporally correlated marker dynamics
- domain assumption Diffusion-based alarm transport combined with leaky gateway evidence integration models the communication channel
Reference graph
Works this paper leans on
-
[1]
Beyond Silicon: Materials, Mechanisms, and Methods for Physical Neural Computing
S. Fischer et al.,Beyond silicon: Materials, mechanisms, and methods for physical neural computing, 2026.DOI: 10.48550/arXiv.2604.09833
work page internal anchor Pith review Pith/arXiv arXiv doi:10.48550/arxiv.2604.09833 2026
-
[2]
DNA-based nanonetwork for abnormality detection and localization in the human body,
J. T. G ´omez et al., “DNA-based nanonetwork for abnormality detection and localization in the human body,”IEEE Transactions on Nanotechnology, vol. 23, pp. 794–808, 2024.DOI: 10.1109/TNANO.2024.3495541
-
[3]
Birkan and Demirkol, Ilker and Farsad, Nariman and Goldsmith, Andrea , title =
M. S ¸. Kuran, H. B. Yilmaz, I. Demirkol, N. Farsad, and A. Goldsmith, “A survey on modulation techniques in molecular communication via diffusion,” IEEE Communications Surveys & Tutorials, vol. 23, no. 1, pp. 7–28, 2021.DOI: 10.1109/COMST.2020.3048099
-
[4]
W. Gao, L. Shi, and L.-L. Yang,Molecular code-division multiple-access: Signaling, detection, and performance, 2024.DOI: 10.48550/arXiv.2402.16097
-
[5]
Nucleic acid strand displacement - from DNA nanotechnology to translational regula- tion,
F. C. Simmel, “Nucleic acid strand displacement - from DNA nanotechnology to translational regulation,”RNA Biol, vol. 20, no. 1, pp. 154–163, Jan. 2023.DOI: 10.1080/15476286.2023.2204565
-
[6]
Scaling up molecular pattern recognition with DNA-based winner-take- all neural networks,
K. M. Cherry and L. Qian, “Scaling up molecular pattern recognition with DNA- based winner-take-all neural networks,”Nature, vol. 559, no. 7714, pp. 370–376, 2018.DOI: 10.1038/s41586-018-0289-6
-
[7]
L. Qian and E. Winfree, “Scaling up digital circuit computation with DNA strand displacement cascades,”Science, vol. 332, no. 6034, pp. 1196–1201, 2011.DOI: 10.1126/science.1200520
-
[8]
Supervised learning in DNA neural networks,
K. M. Cherry and L. Qian, “Supervised learning in DNA neural networks,” Nature, 2025.DOI: 10.1038/s41586-025-09479-w
-
[9]
A comprehensive survey of recent advancements in molecular communication,
N. Farsad, H. B. Yilmaz, A. Eckford, C.-B. Chae, and W. Guo, “A comprehensive survey of recent advancements in molecular communication,”IEEE Communica- tions Surveys & Tutorials, vol. 18, no. 3, pp. 1887–1919, 2016.DOI: 10.1109/ COMST.2016.2527741
-
[10]
Body area nanonetworks with molecular communications in nanomedicine,
B. Atakan, O. B. Akan, and S. Balasubramaniam, “Body area nanonetworks with molecular communications in nanomedicine,”IEEE Communications Magazine, vol. 50, no. 1, pp. 28–34, 2012.DOI: 10.1109/MCOM.2012.6122529
-
[11]
The internet of bio-nano things,
I. F. Akyildiz, M. Pierobon, S. Balasubramaniam, and Y . Koucheryavy, “The internet of bio-nano things,”IEEE Communications Magazine, vol. 53, no. 3, pp. 32–40, 2015.DOI: 10.1109/MCOM.2015.7060516
-
[12]
Modeling and analysis of abnormality detection in biomolecular nano-networks,
S. Ghavami, F. Lahouti, and A. Masoudi-Nejad, “Modeling and analysis of abnormality detection in biomolecular nano-networks,”Nano Communication Networks, vol. 3, no. 4, pp. 229–241, 2012.DOI: 10.1016/j.nancom.2012.09.008
-
[13]
Abnormality detection in correlated gaussian molec- ular nano-networks: Design and analysis,
S. Ghavami and F. Lahouti, “Abnormality detection in correlated gaussian molec- ular nano-networks: Design and analysis,”IEEE Transactions on NanoBioscience, vol. 16, no. 3, pp. 189–202, 2017.DOI: 10.1109/TNB.2017.2659678
-
[14]
Cooperative abnormality detection via diffusive molec- ular communications,
R. Mosayebi, V . Jamali Kooshkghazi, N. Ghoroghchian, R. Schober, M. Nasiri- Kenari, and M. Mehrabi, “Cooperative abnormality detection via diffusive molec- ular communications,”IEEE Transactions on NanoBioscience, vol. 16, no. 8, pp. 828–842, 2017.DOI: 10.1109/TNB.2017.2775704
-
[15]
IEEE Transactions on Molecular, Biological and Multi-Scale Communications , month = Sep, number =
N. Varshney, A. Patel, Y . Deng, W. Haselmayr, P. K. Varshney, and A. Nal- lanathan, “Abnormality detection inside blood vessels with mobile nanoma- chines,”IEEE Transactions on Molecular , Biological, and Multi-Scale Communi- cations, vol. 4, no. 3, pp. 189–194, 2018.DOI: 10.1109/TMBMC.2019.2913399
-
[16]
Using off-the-shelf biosensors to implement gateways for alarm-system nanonetworks,
F.-L. A. Lau et al., “Using off-the-shelf biosensors to implement gateways for alarm-system nanonetworks,”Nano Communication Networks, vol. 45, p. 100 584, Sep. 2025.DOI: 10.1016/j.nancom.2025.100584
-
[17]
Age of information-based abnormality detection with decay in the human circulatory system,
S. Pal, J. Torres G ´omez, R. Wendt, S. Fischer, and F. Dressler, “Age of information-based abnormality detection with decay in the human circulatory system,”IEEE Transactions on Molecular , Biological, and Multi-Scale Commu- nications, vol. 10, no. 3, pp. 487–492, Sep. 2024.DOI: 10.1109/TMBMC.2024. 3426951
-
[18]
DNA strand displacement based computational systems and their applications,
C. Chen, J. Wen, Z. Wen, S. Song, and X. Shi, “DNA strand displacement based computational systems and their applications,”Front Genet, vol. 14, p. 1 120 791, Feb. 2023.DOI: 10.3389/fgene.2023.1120791
-
[19]
Neural net- work computation with DNA strand displacement cascades,
L. Qian, E. Winfree, and J. Bruck, “Neural network computation with DNA strand displacement cascades,”Nature, vol. 475, no. 7356, pp. 368–372, 2011. DOI: 10.1038/nature10262
-
[20]
Deep convolutional and fully-connected DNA neural networks,
X. Liu et al., “Deep convolutional and fully-connected DNA neural networks,” Nat Commun, vol. 16, no. 1, p. 10 629, Nov. 2025.DOI: 10.1038/s41467-025- 65618-x
-
[21]
Nonlinear decision-making with enzymatic neural networks,
S. Okumura et al., “Nonlinear decision-making with enzymatic neural networks,” Nature, vol. 610, no. 7932, pp. 496–501, Oct. 2022.DOI: 10.1038/s41586-022- 05218-7
-
[22]
A molecular multi-gene classifier for disease diagnostics,
R. Lopez, R. Wang, and G. Seelig, “A molecular multi-gene classifier for disease diagnostics,”Nature Chemistry, vol. 10, pp. 746–754, 2018.DOI: 10 . 1038 / s41557-018-0056-1
2018
-
[23]
Cancer diagnosis with DNA molecular computation,
C. Zhang et al., “Cancer diagnosis with DNA molecular computation,”Nature Nanotechnology, vol. 15, no. 8, pp. 709–715, 2020.DOI: 10.1038/s41565-020- 0699-0
-
[24]
A spatially localized DNA linear classifier for cancer diagnosis,
L. Yang et al., “A spatially localized DNA linear classifier for cancer diagnosis,” Nature Communications, vol. 15, p. 4583, 2024.DOI: 10 . 1038 / s41467 - 024 - 48869-y
2024
-
[25]
Calibra- tion and object correspondence in camera networks with widely separated overlapping views,
M. Sanjabi and A. Jahanian, “Multi-threshold and multi-input DNA logic design style for profiling the microrna biomarkers of real cancers,”IET Nanobiotech- nology, vol. 13, no. 7, pp. 665–673, 2019.DOI: https://doi.org/10.1049/iet- nbt.2018.5275
-
[26]
S. Takiguchi et al., “Harnessing DNA computing and nanopore decoding for practical applications: From informatics to microRNA-targeting diagnostics,” Chem. Soc. Rev., vol. 54, pp. 8–32, 1 2025.DOI: 10.1039/D3CS00396E
-
[27]
J. T. G ´omez et al.,Communicating smartly in molecular communication en- vironments: Neural networks in the Internet of Bio-Nano Things, 2025.DOI: 10.48550/arXiv.2506.20589
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.