Recognition: unknown
Cell-cell adhesion cannot sustain extended follower streams in a minimal non-local model of leader-follower migration
Pith reviewed 2026-05-10 09:21 UTC · model grok-4.3
The pith
A minimal non-local adhesion model sustains only short follower cohorts whose length is bounded by the interaction range.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
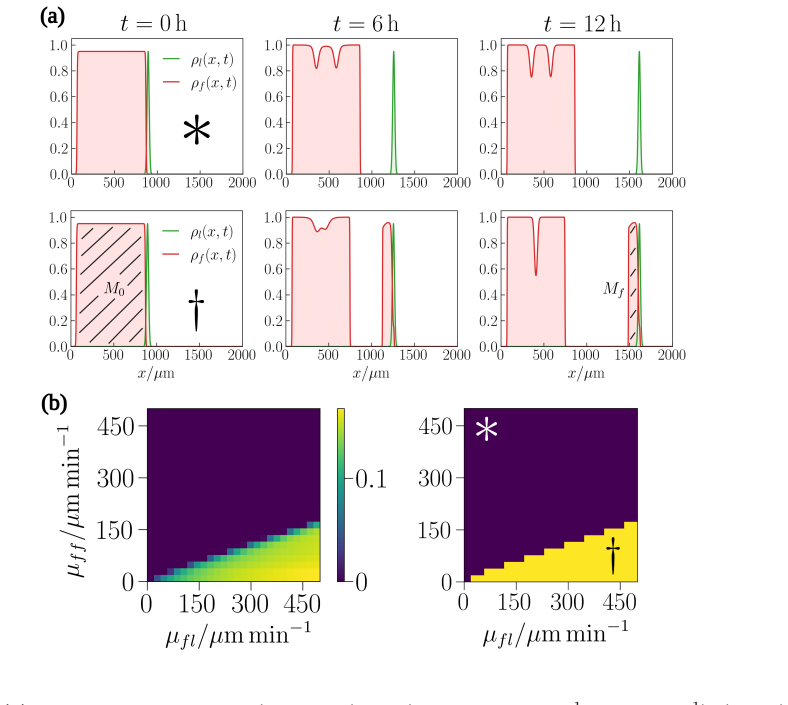
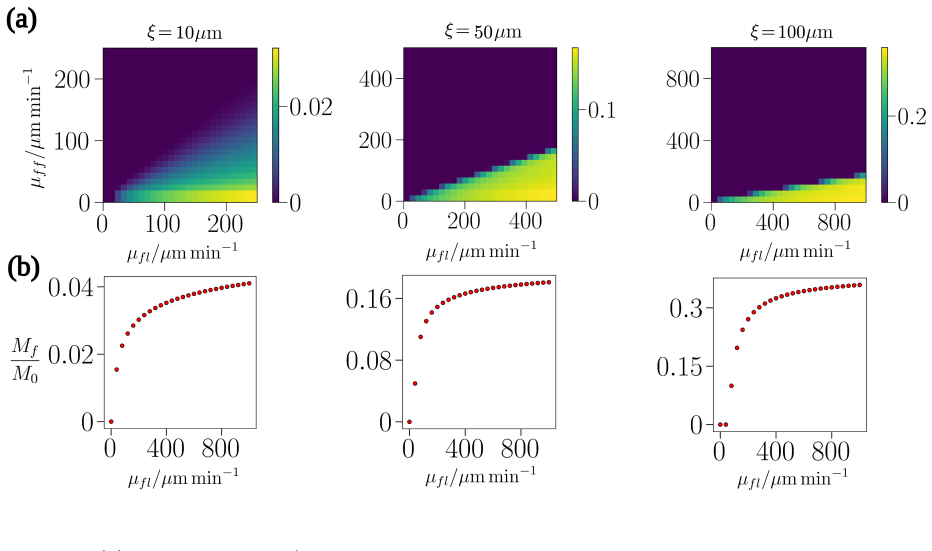
Numerical simulations of the minimal non-local advection-diffusion model reveal that follower cells can form small travelling cohorts behind constant-velocity leaders, but the maximum cohort size is strictly limited by the finite adhesive interaction lengthscale and remains far below the extended streams observed in vivo.
What carries the argument
The minimal non-local advection-diffusion equation for followers, driven by constant-velocity leaders and finite-range attraction kernels between cells.
If this is right
- Small cohorts of followers can travel coherently with leaders through adhesion alone.
- Cohort size cannot exceed the spatial scale of the adhesive interaction.
- Extended, long-distance follower streams cannot arise from mass-conserving non-local adhesion in this framework.
- New continuum models with additional mechanisms are required to reproduce the long migratory streams seen biologically.
Where Pith is reading between the lines
- The limitation may be general to any finite-range non-local attraction term and could persist even with more complex kernels.
- Mechanisms such as cell proliferation, contact inhibition, or long-range chemical signalling may be necessary to overcome the lengthscale bound.
- The result suggests that purely adhesive explanations for stream cohesion need re-examination in other collective migration contexts.
Load-bearing premise
The chosen minimal non-local advection-diffusion framework with constant leader velocity and finite-range attraction represents the essential features of adhesive cell interactions.
What would settle it
A numerical simulation or experiment in which the interaction range is systematically increased while all other parameters remain fixed, and the resulting follower stream length is measured to see whether it grows without bound or stays capped.
Figures

read the original abstract
Cell-cell adhesion is widely hypothesised to maintain cohesion within the long streams of follower cells that trail leader subpopulations during collective migration, including in neural crest cell migration, angiogenesis, and cancer cell invasion. Mathematically, non-local advection-diffusion equations provide the canonical continuum framework within which to study such adhesive cell-cell interactions. Here, we study a minimal model of leader-follower migration within this framework, in which leaders migrate at constant velocity while followers are attracted to leaders and to one another over a finite spatial interaction range. Numerical simulations reveal that, although the model can maintain small cohorts of travelling follower cells, the size of these cohorts is limited by the adhesive interaction lengthscale, and is far below what is needed to reproduce the extended streams observed in vivo. This points to a structural limitation of the standard non-local adhesion formulation and highlights the need for the development of extended continuum models capable of sustaining long, coherent migratory streams through purely mass-conserving collective cell movement.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript studies a minimal non-local advection-diffusion model of leader-follower cell migration in which leaders advance at constant velocity while followers experience finite-range attraction to both leaders and other followers. Numerical simulations show that stable travelling follower cohorts form but remain bounded in size by the adhesive interaction lengthscale, with maximum sizes far smaller than the extended streams observed in vivo (e.g., neural crest, angiogenesis). The authors interpret this as evidence of a structural limitation in the standard continuum formulation of cell-cell adhesion and call for extended models that can sustain long coherent streams under mass conservation.
Significance. If the reported scaling holds, the result is significant because it supplies a clear negative finding for the canonical minimal non-local adhesion model that is widely used to explain collective migration. By restricting to constant leader speed and finite-range interactions, the work isolates the limitation without additional mechanisms, thereby directing future modelling efforts toward non-local kernels with longer effective range, dynamic adhesion, or additional transport terms. The explicit framing as a limitation of the minimal setup rather than a universal claim increases the utility of the negative result for the field.
minor comments (3)
- [Numerical methods] The numerical methods section should include a brief convergence study (grid size, time step, and interaction kernel discretisation) to confirm that the reported cohort-size bound is not an artifact of the discretisation scheme.
- [Results] A table listing all parameter values (including the range of adhesive interaction lengths tested and the corresponding steady-state cohort sizes) would improve reproducibility and allow readers to judge how far below in-vivo lengths the model cohorts remain.
- [Discussion] The discussion could briefly contrast the finite-range kernel used here with alternative non-local formulations (e.g., those with decaying but infinite-range kernels) to clarify why the size limitation is specific to the standard finite-range choice.
Simulated Author's Rebuttal
We thank the referee for their positive assessment of the manuscript and their recommendation to accept. The referee's summary accurately reflects the central result: that the minimal non-local adhesion model produces only bounded follower cohorts whose size is limited by the interaction lengthscale, far below the extended streams seen in vivo.
Circularity Check
No significant circularity detected
full rationale
The paper defines a minimal non-local advection-diffusion model with constant leader velocity and finite-range attraction, then reports direct numerical simulation outcomes showing that follower cohort sizes are bounded by the interaction lengthscale. This negative result is obtained by integrating the stated PDE system and observing the emergent scaling; it does not reduce to a fitted parameter renamed as a prediction, a self-definitional loop, or any load-bearing self-citation. The authors explicitly frame the work as exposing a limitation of the standard formulation rather than claiming a first-principles derivation that collapses to its inputs.
Axiom & Free-Parameter Ledger
free parameters (2)
- adhesive interaction lengthscale
- leader velocity
axioms (2)
- standard math Mass conservation in the advection-diffusion system
- domain assumption Finite-range non-local attraction between followers and leaders
Reference graph
Works this paper leans on
-
[1]
Collective action in birds.Current Biology, 32(20):R1140–R1144, 2022
Damien R Farine. Collective action in birds.Current Biology, 32(20):R1140–R1144, 2022
2022
-
[2]
From behavioural analyses to models of collective motion in fish schools.Interface Focus, 2(6):693–707, 2012
Ugo Lopez, Jacques Gautrais, Iain D Couzin, and Guy Theraulaz. From behavioural analyses to models of collective motion in fish schools.Interface Focus, 2(6):693–707, 2012
2012
-
[3]
Collective cell migration.Annual Review of Cell and Developmental Biology, 25:407–429, 2009
Pernille Rørth. Collective cell migration.Annual Review of Cell and Developmental Biology, 25:407–429, 2009
2009
-
[4]
Collective cell migration in development.Journal of Cell Biology, 212(2):143–155, 2016
Elena Scarpa and Roberto Mayor. Collective cell migration in development.Journal of Cell Biology, 212(2):143–155, 2016. 12
2016
-
[5]
Collective cell migration: Implications for wound healing and cancer invasion.Burns & Trauma, 1(1):21–26, 2013
Li Li, Yong He, Min Zhao, and Jianxin Jiang. Collective cell migration: Implications for wound healing and cancer invasion.Burns & Trauma, 1(1):21–26, 2013
2013
-
[6]
Collective cell migration in morphogenesis and cancer
Peter Friedl, Yael Hegerfeldt, and Miriam Tusch. Collective cell migration in morphogenesis and cancer. International Journal of Developmental Biology, 48(5-6):441–449, 2004
2004
-
[7]
The principles of directed cell migration
Shuvasree SenGupta, Carole A Parent, and James E Bear. The principles of directed cell migration. Nature Reviews Molecular Cell Biology, 22(8):529–547, 2021
2021
-
[8]
Cell interactions in collective cell migration.Development, 146(23):dev172056, 2019
Abhinava K Mishra, Joseph P Campanale, James A Mondo, and Denise J Montell. Cell interactions in collective cell migration.Development, 146(23):dev172056, 2019
2019
-
[9]
Cranial neural crest migration: new rules for an old road.Developmental Biology, 344(2):543–554, 2010
Paul M Kulesa, Caleb M Bailey, Jennifer C Kasemeier-Kulesa, and Rebecca McLennan. Cranial neural crest migration: new rules for an old road.Developmental Biology, 344(2):543–554, 2010
2010
-
[10]
Biological waves: single species models
James D Murray. Biological waves: single species models. InMathematical Biology, pages 437–482. Springer, 2004
2004
-
[11]
Matthew J Simpson and Scott W McCue. Fisher–KPP-type models of biological invasion: open source computational tools, key concepts and analysis.Proceedings of the Royal Society A: Mathematical, Physical and Engineering Sciences, 480(2294), 2024
2024
-
[12]
Looking inside an invasion wave of cells using continuum models: proliferation is the key.Journal of Theoretical Biology, 243(3):343–360, 2006
Matthew J Simpson, Kerry A Landman, Barry D Hughes, and Donald F Newgreen. Looking inside an invasion wave of cells using continuum models: proliferation is the key.Journal of Theoretical Biology, 243(3):343–360, 2006
2006
-
[13]
Cell proliferation drives neural crest cell invasion of the intestine.Developmental Biology, 302(2):553–568, 2007
Matthew J Simpson, Dong C Zhang, Michael Mariani, Kerry A Landman, and Donald F Newgreen. Cell proliferation drives neural crest cell invasion of the intestine.Developmental Biology, 302(2):553–568, 2007
2007
-
[14]
The neural crest cell cycle is related to phases of migration in the head.Development, 141(5):1095–1103, 2014
Dennis A Ridenour, Rebecca McLennan, Jessica M Teddy, Craig L Semerad, Jeffrey S Haug, and Paul M Kulesa. The neural crest cell cycle is related to phases of migration in the head.Development, 141(5):1095–1103, 2014. 13
2014
-
[15]
Self- generated chemotaxis of mixed cell populations.Proceedings of the National Academy of Sciences, 122(34):e2504064122, 2025
Mehmet Can U¸ car, Zane Alsberga, Jonna Alanko, Michael Sixt, and Edouard Hannezo. Self- generated chemotaxis of mixed cell populations.Proceedings of the National Academy of Sciences, 122(34):e2504064122, 2025
2025
-
[16]
A continuum approach to modelling cell–cell adhesion.Journal of Theoretical Biology, 243(1):98–113, 2006
Nicola J Armstrong, Kevin J Painter, and Jonathan A Sherratt. A continuum approach to modelling cell–cell adhesion.Journal of Theoretical Biology, 243(1):98–113, 2006
2006
-
[17]
A population dynamics model of cell-cell adhesion incorporating population pressure and density saturation.Journal of Theoretical Biology, 474:14–24, 2019
Jose A Carrillo, Hideki Murakawa, Makoto Sato, Hideru Togashi, and Olena Trush. A population dynamics model of cell-cell adhesion incorporating population pressure and density saturation.Journal of Theoretical Biology, 474:14–24, 2019
2019
-
[18]
Biological modeling with nonlocal advection– diffusion equations.Mathematical Models and Methods in Applied Sciences, 34(1):57–107, 2024
Kevin J Painter, Thomas Hillen, and Jonathan R Potts. Biological modeling with nonlocal advection– diffusion equations.Mathematical Models and Methods in Applied Sciences, 34(1):57–107, 2024
2024
-
[19]
Valeria Giunta, Thomas Hillen, Mark A Lewis, and Jonathan R Potts. A phylogeny of biological patterns formed by nonlocal advection.arXiv preprint arXiv:2506.00489, 2025
-
[20]
Synergistic action of nectins and cadherins generates the mosaic cellular pattern of the olfactory epithelium.Journal of Cell Biology, 212(5):561–575, 2016
Sayaka Katsunuma, Hisao Honda, Tomoyasu Shinoda, Yukitaka Ishimoto, Takaki Miyata, Hiroshi Kiy- onari, Takaya Abe, Ken-ichi Nibu, Yoshimi Takai, and Hideru Togashi. Synergistic action of nectins and cadherins generates the mosaic cellular pattern of the olfactory epithelium.Journal of Cell Biology, 212(5):561–575, 2016
2016
-
[21]
Directed cell migration is a versatile mechanism for rapid developmental pattern formation.bioRxiv, 2025
Chengyou Yu, Malte Mederacke, Roman Vetter, and Dagmar Iber. Directed cell migration is a versatile mechanism for rapid developmental pattern formation.bioRxiv, 2025
2025
-
[22]
Patterning of nonlocal transport models in biology: the impact of spatial dimension.Mathematical Biosciences, 366:109093, 2023
Thomas Jun Jewell, Andrew L Krause, Philip K Maini, and Eamonn A Gaffney. Patterning of nonlocal transport models in biology: the impact of spatial dimension.Mathematical Biosciences, 366:109093, 2023
2023
-
[23]
A nonlocal model for contact attraction and repulsion in heterogeneous cell populations.Bulletin of Mathematical Biology, 77(6):1132–1165, 2015
Kevin J Painter, Jennifer M Bloomfield, Jonathan A Sherratt, and Alf Gerisch. A nonlocal model for contact attraction and repulsion in heterogeneous cell populations.Bulletin of Mathematical Biology, 77(6):1132–1165, 2015. 14
2015
-
[24]
Mathematical modelling of cancer invasion: implications of cell adhesion variability for tumour infiltrative growth patterns.Journal of Theoretical Biology, 361:41–60, 2014
Pia Domschke, Dumitru Trucu, Alf Gerisch, and Mark AJ Chaplain. Mathematical modelling of cancer invasion: implications of cell adhesion variability for tumour infiltrative growth patterns.Journal of Theoretical Biology, 361:41–60, 2014
2014
-
[25]
Mathematical modelling of cancer cell invasion of tissue: local and non-local models and the effect of adhesion.Journal of Theoretical Biology, 250(4):684–704, 2008
Alf Gerisch and Mark AJ Chaplain. Mathematical modelling of cancer cell invasion of tissue: local and non-local models and the effect of adhesion.Journal of Theoretical Biology, 250(4):684–704, 2008
2008
-
[26]
The impact of adhesion on cellular invasion processes in cancer and development.Journal of Theoretical Biology, 264(3):1057–1067, 2010
Kevin J Painter, Nicola J Armstrong, and Jonathan A Sherratt. The impact of adhesion on cellular invasion processes in cancer and development.Journal of Theoretical Biology, 264(3):1057–1067, 2010
2010
-
[27]
Chiara Villa, Alf Gerisch, and Mark AJ Chaplain. A novel nonlocal partial differential equation model of endothelial progenitor cell cluster formation during the early stages of vasculogenesis.Journal of Theoretical Biology, 534:110963, 2022
2022
-
[28]
Modeling the morphodynamic galectin pat- terning network of the developing avian limb skeleton.Journal of Theoretical Biology, 346:86–108, 2014
Tilmann Glimm, Ramray Bhat, and Stuart A Newman. Modeling the morphodynamic galectin pat- terning network of the developing avian limb skeleton.Journal of Theoretical Biology, 346:86–108, 2014
2014
-
[29]
Distinguishing between long-transient and asymptotic states in a biological aggregation model.Bulletin of Mathematical Biology, 86(3):28, 2024
Jonathan R Potts and Kevin J Painter. Distinguishing between long-transient and asymptotic states in a biological aggregation model.Bulletin of Mathematical Biology, 86(3):28, 2024
2024
-
[30]
Thomas Jun Jewell, Andrew L Krause, Philip K Maini, and Eamonn A Gaffney. Chase-and-run and chirality in nonlocal models of pattern formation.arXiv preprint arXiv:2505.17372, 2025
-
[31]
Zoology of a nonlocal cross-diffusion model for two species.SIAM Journal on Applied Mathematics, 78(2):1078–1104, 2018
Jos´ e A Carrillo, Yanghong Huang, and Markus Schmidtchen. Zoology of a nonlocal cross-diffusion model for two species.SIAM Journal on Applied Mathematics, 78(2):1078–1104, 2018
2018
-
[32]
Variations in non-local interac- tion range lead to emergent chase-and-run in heterogeneous populations.Journal of the Royal Society Interface, 21(219):20240409, 2024
Kevin J Painter, Valeria Giunta, Jonathan R Potts, and Sara Bernardi. Variations in non-local interac- tion range lead to emergent chase-and-run in heterogeneous populations.Journal of the Royal Society Interface, 21(219):20240409, 2024
2024
-
[33]
Aggregation–diffusion in heterogeneous environments.Journal of Mathematical Biology, 90(6):59, 2025
Jonathan R Potts. Aggregation–diffusion in heterogeneous environments.Journal of Mathematical Biology, 90(6):59, 2025. 15
2025
-
[34]
The impact of different degrees of leadership on collective navigation in follower-leader systems.Bulletin of Mathematical Biology, 87(5):1–31, 2025
Sara Bernardi and Kevin J Painter. The impact of different degrees of leadership on collective navigation in follower-leader systems.Bulletin of Mathematical Biology, 87(5):1–31, 2025. 16 Appendix A: Model Formulation and Numerical Scheme A.1 Numerical scheme We discretise the domain intoN= 10 4 mesh points,{x i}N i=1 with uniform spacing,h=L/(N−1). The s...
2025
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.