Recognition: unknown
Universal Loop Statistics from Active Extrusion with Kinetic Barriers
Pith reviewed 2026-05-10 09:18 UTC · model grok-4.3
The pith
Mean chromatin loop sizes obey a universal law set by processivity and renormalized barrier density, while length distributions distinguish one-sided from two-sided extrusion.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
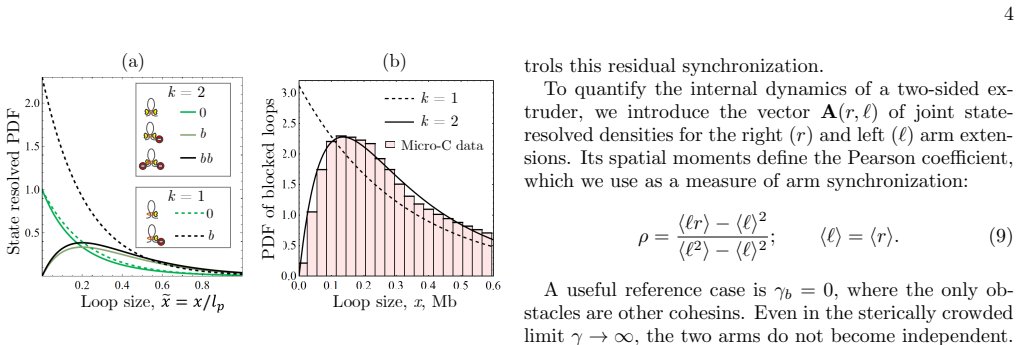
In the stationary state the mean loop size obeys a universal law determined by the bare processivity and a renormalized obstacle density. One-sided extrusion always yields a single-exponential loop-length distribution, whereas two-sided extrusion produces a finite sum of exponential modes and, generically, a peaked distribution. Experimental CTCF-anchored loop statistics exhibit such a peak, thereby providing a direct discriminator of extrusion symmetry and supporting a scenario in which both cohesin arms actively operate in living cells.
What carries the argument
Kinetic theory of active extrusion on a disordered chromatin track with transient barriers, whose effects are captured by a renormalized obstacle density that controls stationary loop statistics.
Load-bearing premise
The model assumes the system reaches a stationary state in which barrier effects reduce to a simple renormalized density without requiring their detailed time-dependent dynamics.
What would settle it
Measuring the shape of CTCF-anchored loop-length distributions in high-resolution experiments and checking whether the distribution is peaked (two-sided) or purely exponential (one-sided) would directly test the central prediction.
Figures



read the original abstract
We develop a kinetic theory of cohesin-driven loop extrusion on a disordered chromatin track with transient barriers. In the stationary state, the mean loop size is shown to obey a universal law determined by the bare processivity and a renormalized obstacle density. Beyond the mean, one-sided extrusion always yields a single-exponential loop-length distribution, whereas two-sided extrusion produces a finite sum of exponential modes and, generically, a peaked distribution. Experimental CTCF-anchored loop statistics exhibit such a peak, thereby providing a direct discriminator of extrusion symmetry. The theory therefore establishes a unified framework for disorder-limited loop extrusion and supports a scenario in which both cohesin arms actively operate in living cells.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript develops a kinetic theory of cohesin-driven loop extrusion on a disordered chromatin track with transient barriers. In the stationary state, the mean loop size is shown to obey a universal law determined by the bare processivity and a renormalized obstacle density. One-sided extrusion always yields a single-exponential loop-length distribution, whereas two-sided extrusion produces a finite sum of exponential modes and generically a peaked distribution. Experimental CTCF-anchored loop statistics exhibit such a peak, providing a discriminator of extrusion symmetry and supporting two-sided active extrusion in cells.
Significance. If the derivations hold, the work supplies a unified framework for disorder-limited loop extrusion with a clear, testable distinction between extrusion modes via loop-length distributions. The universal mean-size law and the one- versus two-sided shape predictions are potentially impactful for chromatin biology and Hi-C data interpretation, especially if the renormalization step is shown to be robust.
major comments (2)
- [stationary state analysis] The central claim that the mean loop size obeys a universal law determined solely by bare processivity and renormalized obstacle density (abstract and stationary-state analysis) rests on the renormalization of transient barriers collapsing to a single density parameter. No explicit demonstration is provided that this renormalization is independent of barrier on/off rates, spatial correlations, or loading/unloading details; if barrier kinetics introduce additional timescales, the stationary distribution acquires extra parameters and universality is lost.
- [distribution derivations] The assertion that two-sided extrusion generically produces a peaked distribution (as an experimental discriminator) depends on the same renormalization step remaining parameter-free. If the finite sum of exponential modes is sensitive to the unspecified barrier dynamics, the claimed distinction between one-sided and two-sided cases is undermined.
minor comments (3)
- Provide explicit equations for the renormalization procedure and the resulting mean loop size formula at the earliest point in the theory section.
- Include more detail on simulation protocols, error analysis, and how experimental CTCF data were processed and compared to the theoretical distributions.
- Clarify the definition of 'bare processivity' with its precise relation to the microscopic rates.
Simulated Author's Rebuttal
We thank the referee for their careful reading of the manuscript and for the constructive comments. We appreciate the recognition of the potential impact of the unified framework and the distinction between extrusion modes. We address the major comments point by point below, clarifying the derivations and indicating revisions to strengthen the presentation of the renormalization step.
read point-by-point responses
-
Referee: The central claim that the mean loop size obeys a universal law determined solely by bare processivity and renormalized obstacle density (abstract and stationary-state analysis) rests on the renormalization of transient barriers collapsing to a single density parameter. No explicit demonstration is provided that this renormalization is independent of barrier on/off rates, spatial correlations, or loading/unloading details; if barrier kinetics introduce additional timescales, the stationary distribution acquires extra parameters and universality is lost.
Authors: We thank the referee for highlighting this important point regarding the robustness of the renormalization. In the stationary-state analysis, the master equations for loop growth and barrier encounters are solved by averaging the transient barrier occupancy, yielding an effective obstacle density that enters the mean loop size as a single parameter alongside the bare processivity. The derivation assumes independent barriers and incorporates on/off rates into this effective density. However, we acknowledge that an explicit verification of independence from specific rate values (while holding the renormalized density fixed) was not provided. We will add an appendix with analytic limits and numerical solutions of the full kinetic model across varied on/off rates and loading details, confirming collapse to the universal law. This revision will be incorporated. revision: yes
-
Referee: The assertion that two-sided extrusion generically produces a peaked distribution (as an experimental discriminator) depends on the same renormalization step remaining parameter-free. If the finite sum of exponential modes is sensitive to the unspecified barrier dynamics, the claimed distinction between one-sided and two-sided cases is undermined.
Authors: The loop-length distribution for two-sided extrusion is obtained by solving the coupled steady-state equations for the two arms, resulting in a finite sum of exponential modes whose decay rates and amplitudes are set by the effective parameters after renormalization. One-sided extrusion reduces to a single exponential by construction. Because the renormalization reduces barrier kinetics to the single effective density (whose independence will be demonstrated in the new appendix), the generic presence of a peak for two-sided extrusion and the distinction from the one-sided case remain intact. We will revise the relevant sections to explicitly note this dependence on the renormalized parameters and add a short robustness discussion. This addresses the concern. revision: yes
Circularity Check
No circularity: kinetic derivation of universal loop statistics is self-contained
full rationale
The paper presents a kinetic theory deriving stationary-state loop statistics from processivity and renormalized obstacle density. The mean loop size law and the one-sided vs. two-sided distribution distinctions follow from solving the model equations under the stated assumptions of stationary state and effective barrier renormalization. No quoted step reduces the target result to a fitted parameter or self-citation by construction; renormalization is introduced as a modeling approximation rather than a tautological redefinition of the mean. The experimental discriminator is a model prediction, not a post-hoc fit. This is a standard non-circular theoretical derivation.
Axiom & Free-Parameter Ledger
free parameters (2)
- bare processivity
- renormalized obstacle density
axioms (2)
- domain assumption The system reaches a stationary state
- domain assumption Barriers are transient kinetic obstacles on a disordered track
Reference graph
Works this paper leans on
-
[1]
Dekker and L
J. Dekker and L. Mirny, Cell187, 6424 (2024)
2024
-
[2]
Fraser and W
P. Fraser and W. Bickmore, Nature447, 413 (2007)
2007
-
[3]
A. L. Sanborn, S. S. Rao, S. C. Huang, N. C. Durand, M. H. Huntley, A. I. Jewett,et al., Proc. Natl. Acad. Sci. U.S.A.112, E6456 (2015)
2015
-
[4]
Fudenberg, M
G. Fudenberg, M. Imakaev, C. Lu, A. Goloborodko, N. Abdennur, and L. Mirny, Cell Rep.15, 2038 (2016)
2038
-
[5]
C. A. Brackley, J. Johnson, D. Michieletto, A. N. Mo- rozov, M. Nicodemi, P. R. Cook, and D. Marenduzzo, Phys. Rev. Lett.119, 138101 (2017)
2017
-
[6]
S. S. Rao, S. C. Huang, B. G. St Hilaire, J. M. Engreitz, E. M. Perez,et al., Cell171, 305 (2017)
2017
-
[7]
Machet al., Nat
P. Machet al., Nat. Genet.54, 1907 (2022)
1907
-
[8]
Gabriele, H
M. Gabriele, H. B. Brand˜ ao, S. Grosse-Holz, A. Jha, G. M. Dailey, C. Cattoglio,et al., Science376, 496 (2022)
2022
-
[9]
Sabat´ eet al., Nat
T. Sabat´ eet al., Nat. Genet.57, 3152–3164 (2025)
2025
-
[10]
H. B. Brand˜ ao, P. Paul, A. A. van den Berg, D. Z. Rud- ner, X. Wang, and L. A. Mirny, Proc. Natl. Acad. Sci. U.S.A.116, 20489 (2019)
2019
-
[11]
B. J. Dequekeret al., Nature606, 197 (2022)
2022
-
[12]
Metzler and J
R. Metzler and J. Klafter, Phys. Rep.339, 1 (2000)
2000
-
[13]
Goychuk and V
I. Goychuk and V. O. Kharchenko, Phys. Rev. Lett.113, 100601 (2014)
2014
-
[14]
Goychuk, V
I. Goychuk, V. O. Kharchenko, and R. Metzler, Phys. Chem. Chem. Phys.16, 16524 (2024)
2024
-
[15]
D. S. Fisher, P. Le Doussal, and C. Monthus, Phys. Rev. Lett.80, 3539 (1998)
1998
-
[16]
Burov and E
S. Burov and E. Barkai, Phys. Rev. Lett.98, 250601 (2007)
2007
-
[17]
Y. G. Sinai, Theory Probab. Appl.27, 256 (1983)
1983
-
[18]
J. P. Bouchaud and A. Georges, Phys. Rep.195, 127 6 (1990)
1990
-
[19]
Padash, E
A. Padash, E. Aghion, A. Schulz, E. Barkai, A. V. Chechkin, R. Metzler, and H. Kantz, New J. Phys.24, 073026 (2022)
2022
-
[20]
Oshanin, A
G. Oshanin, A. Rosso, and G. Schehr, Phys. Rev. Lett. 110, 100602 (2013)
2013
-
[21]
Derrida, Phys
B. Derrida, Phys. Rep.301, 65 (1998)
1998
-
[22]
M. R. Evans, Braz. J. Phys.30, 42 (2000)
2000
-
[23]
T. Chou, K. Mallick, and R. K. Zia, Rep. Prog. Phys. 74, 116601 (2011)
2011
-
[24]
I. F. Davidson, B. Bauer, D. Goetz, W. Tang, G. Wutz, and J. M. Peters, Science366, 1338 (2019)
2019
-
[25]
Y. Kim, Z. Shi, H. Zhang, I. J. Finkelstein, and H. Yu, Science366, 1345 (2019)
2019
-
[26]
Ganjiet al., Science360, 102 (2018)
M. Ganjiet al., Science360, 102 (2018)
2018
-
[27]
Nuebler, G
J. Nuebler, G. Fudenberg, M. Imakaev, N. Abdennur, and L. Mirny, Proc. Natl. Acad. Sci. U.S.A.115, E6697 (2018)
2018
-
[28]
Lizana and Y
L. Lizana and Y. B. Schwartz, Sci. Adv.15, eadm8167 (2024)
2024
-
[29]
Bonato, M
A. Bonato, M. Chiang, D. Corbett, S. Kitaev, D. Maren- duzzo, A. Morozov, and E. Orlandini, Phys. Rev. Lett. 132, 248403 (2024)
2024
-
[30]
Forte, S
G. Forte, S. Buonomo, P. R. Cook, N. Gilbert, D. Maren- duzzo, and E. Orlandini, PRX Life2, 033014 (2024)
2024
-
[31]
Di Stefano and G
M. Di Stefano and G. Cavalli, Curr. Opin. Struct. Biol. 77, 102493 (2022)
2022
-
[32]
S. K. Nomidis, E. Carlon, S. Gruber, and J. F. Marko, Nucleic Acids Res.50, 4974 (2022)
2022
-
[33]
Takaki, A
R. Takaki, A. Dey, G. Shi, and D. Thirumalai, Nat. Com- mun.12, 5865 (2021)
2021
-
[34]
M. M. Tortora and G. Fudenberg, Cell Genom.6, 101098 (2026)
2026
-
[35]
Salari and D
H. Salari and D. Jost, Phys. Rev. Research8, 013188 (2026)
2026
-
[36]
E. J. Banigan, A. A. van den Berg, H. B. Brand˜ ao, J. F. Marko, and L. A. Mirny, eLife9, e53558 (2020)
2020
-
[37]
Chan and M
B. Chan and M. Rubinstein, Proc. Natl. Acad. Sci. U.S.A.120, e2222078120 (2023)
2023
-
[38]
A. J. Pinto, B. Pradhan, D. Tetiker, M. P. Schmitt, E. Kim, and P. Virnau, bioRxiv: https://doi.org/10.64898/2026.03.05.709531 (2026)
-
[39]
Chan and M
B. Chan and M. Rubinstein, Proc. Natl. Acad. Sci. U.S.A.121, e2401494121 (2024)
2024
-
[40]
Barthet al., Cell188, 749 (2025)
R. Barthet al., Cell188, 749 (2025)
2025
-
[41]
Oldenkamp and B
R. Oldenkamp and B. D. Rowland, Mol. Cell82, 1616 (2022)
2022
-
[42]
Polovnikov and D
K. Polovnikov and D. Starkov, Proc. Natl. Acad. Sci. U.S.A.123, e2534385123 (2026)
2026
-
[43]
K. E. Polovnikov, H. B. Brand˜ ao, S. Belan, B. Slavov, M. Imakaev, and L. A. Mirny, Phys. Rev. X13, 041029 (2023)
2023
-
[44]
K. E. Polovnikov and B. Slavov, Phys. Rev. E107, 054135 (2023)
2023
-
[45]
Slavov and K
B. Slavov and K. Polovnikov, JETP Lett.118, 208 (2023)
2023
-
[46]
Belan and V
S. Belan and V. Parfenyev, J. Chem. Phys.160, 124901 (2024)
2024
-
[47]
If a roadblock stops cohesin with probabilityp i upon en- counter, this partial permeability is absorbed into the effective spacing, which exceeds the raw genomic spacing by a factorp −1 i
-
[48]
See Supplemental Material at [URL will be inserted by publisher] for details
-
[49]
C. E. Sept, Y. E. Tak, V. Goel, M. S. Bhakta, C. G. Cerda-Smith, H. M. Hutchinson, M. Blanchette, C. E. Eyler, S. E. Johnstone, J. K. Joung,et al., Nat. Commun. 16, 4506 (2025)
2025
-
[50]
E. J. Banigan and L. A. Mirny, Phys. Rev. X9, 031007 (2019)
2019
-
[51]
Brunner, N
A. Brunner, N. R. Morero, W. Zhang, M. J. Hos- sain, M. Lampe, H. Pflaumer,et al., J. Cell Biol.224, e202405169 (2025)
2025
-
[52]
Holzmann, A
J. Holzmann, A. Z. Politi, K. Nagasaka, M. Hantsche- Grininger, N. Walther, B. Koch,et al., eLife8, e46269 (2019)
2019
-
[53]
I. F. Davidson, R. Barth, M. Zaczek, J. van der Torre, W. Tang, K. Nagasaka, R. Janissen, J. Kerssemakers, G. Wutz, C. Dekker,et al., Nature616, 822 (2023)
2023
-
[54]
N. A. Kulak, G. Pichler, I. Paron, N. Nagaraj, and M. Mann, Nat. Methods11, 319 (2014)
2014
-
[55]
Ubertiniet al., bioRxiv, doi: https://doi.org/10.1101/2025.09.24.678119 (2025)
M. Ubertiniet al., bioRxiv, doi: https://doi.org/10.1101/2025.09.24.678119 (2025)
-
[56]
Arnouldet al., Nature590, 660 (2021)
C. Arnouldet al., Nature590, 660 (2021)
2021
-
[57]
J. H. Yang, H. B. Brand˜ ao, and A. S. Hansen, Nat. Com- mun.14, 1913 (2023)
1913
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.