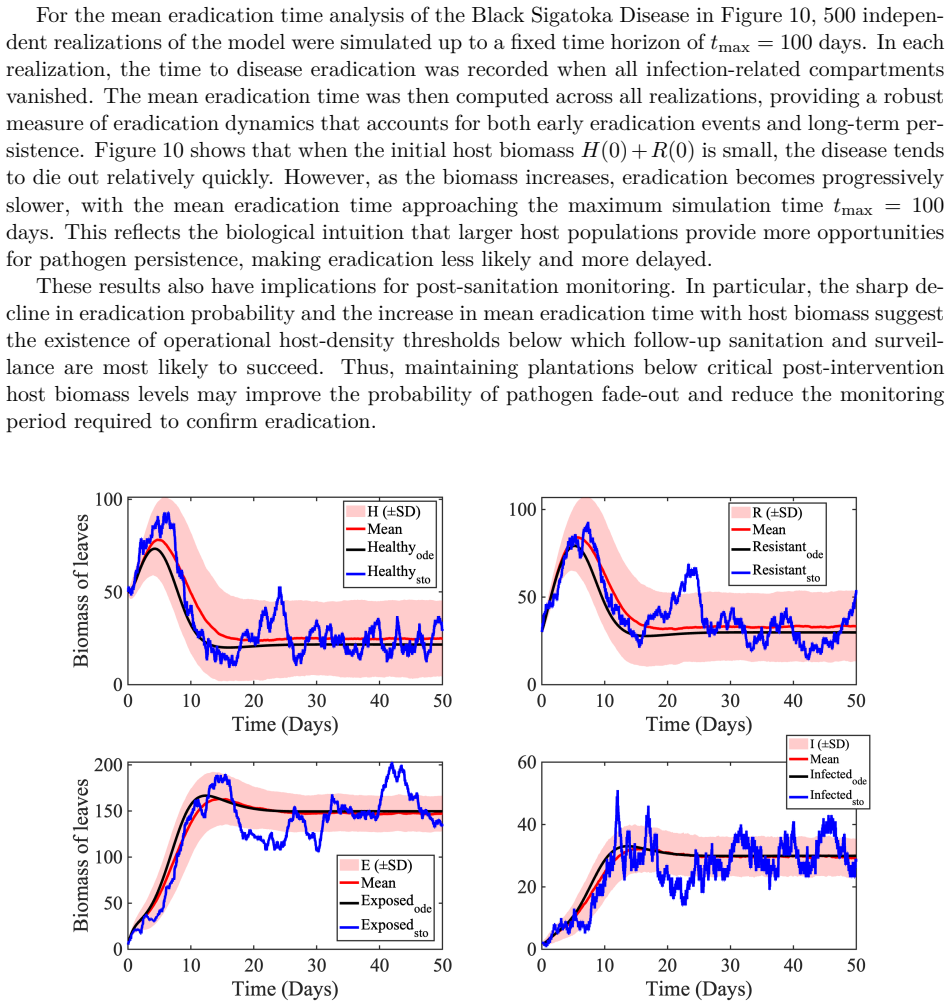
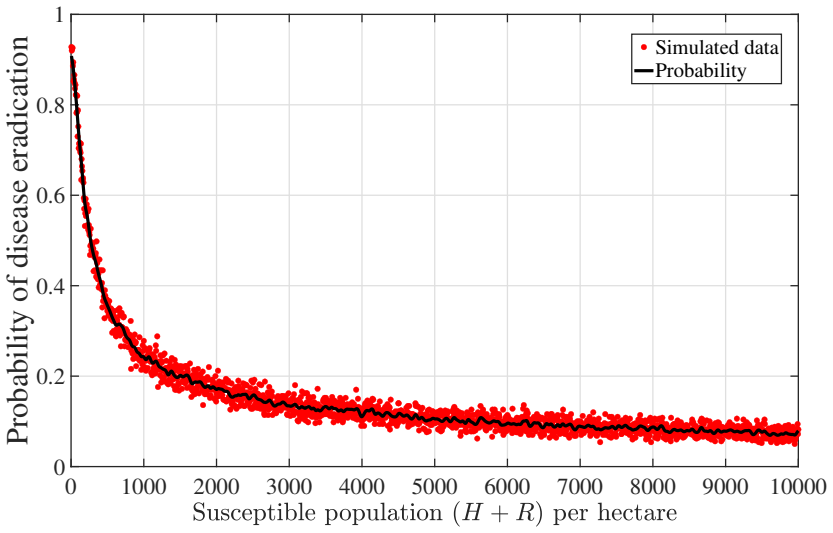
Recognition: unknown
The Curse of Black Sigatoka: A Backward Bifurcation Perspective
Pith reviewed 2026-05-07 13:42 UTC · model grok-4.3
The pith
Black Sigatoka can maintain a stable presence in banana plants even when the basic reproduction number falls below one.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
The deterministic pathogen-host model exhibits backward bifurcation arising from dual transmission pathways and mate limitation in sexual reproduction. As a result a stable endemic equilibrium coexists with the disease-free equilibrium for certain parameter values in which the basic reproduction number is less than one. This explains why control strategies that act only by reducing the reproduction number below one may fail to eradicate the disease. Sensitivity analysis of the endemic equilibrium and stochastic simulations with the Gillespie algorithm further show that limiting production of new susceptible leaves and deploying resistant varieties are key additional controls.
What carries the argument
Backward bifurcation in the compartmental model, produced by the combination of dual spore transmission routes and mate limitation in sexual reproduction.
If this is right
- Reducing only the basic reproduction number below one does not guarantee elimination of the endemic state.
- Effective management must also limit production of new susceptible leaves during high-risk periods.
- Development and deployment of disease-resistant plant varieties becomes a necessary control measure.
- Stochastic simulations indicate that nonlinear parameter interactions drive additional variability in disease persistence.
Where Pith is reading between the lines
- Models of other fungal diseases that use multiple spore types and sexual reproduction may exhibit the same backward bifurcation behavior.
- Controlled field trials that measure whether the disease persists after measured reproduction numbers are driven below one would directly test the predicted coexistence.
- Adding seasonal or climate-driven variation in leaf production to the model could shift the parameter region where backward bifurcation occurs.
Load-bearing premise
The chosen functional forms for dual transmission and the assumption of mate limitation in sexual reproduction accurately reflect the fungus biology; if they do not, the coexistence of a stable endemic equilibrium when the reproduction number is less than one would not occur.
What would settle it
Field data from banana plantations that show the disease is fully eradicated in every case where interventions bring the estimated basic reproduction number below one.
Figures








read the original abstract
Black Sigatoka disease (BSD), also known as black leaf streak disease, is an airborne fungal infection caused by \textit{Pseudocercospora fijiensis} that severely impacts global banana and plantain production. Its persistence and resistance to eradication make it one of the most challenging plant diseases to manage. In this paper, we propose a deterministic pathogen-host model to describe BSD dynamics. Due to dual transmission pathways (ascospores and conidia) and mate limitation in sexual reproduction, the model exhibits a backward bifurcation: a stable endemic equilibrium coexists with the disease-free equilibrium for certain parameter values in which the basic reproduction number, $\mathcal{R}_0$, is less than 1. This phenomenon explains why control strategies that solely reduce $\mathcal{R}_0$ below one may fail. For the backward bifurcation regime, we perform sensitivity analysis of the endemic equilibrium using normalized forward sensitivity indices, Latin Hypercube Sampling, and Partial Rank Correlation Coefficients. Results indicate that effective control must extend beyond $\mathcal{R}_0$ reduction and prioritize (1) limiting production of new susceptible leaves during high-risk periods and (2) developing and deploying disease-resistant plant varieties. To incorporate transmission variability, we also formulate a stochastic version of the model using the Stochastic Simulation Algorithm (SSA). Extensive numerical simulations compare stochastic realizations with deterministic predictions and quantify variability in disease dynamics. To identify the principal drivers of persistence and variability, we analyze the endemic equilibrium using Sobol's variance-based sensitivity method, which highlights the role of nonlinear parameter interactions in shaping variability.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper proposes a deterministic compartmental model for Black Sigatoka disease incorporating dual transmission via ascospores and conidia together with mate limitation in sexual reproduction of the pathogen. It claims that this structure produces a backward bifurcation, so that a stable endemic equilibrium coexists with the disease-free equilibrium for some parameter values where the basic reproduction number R0 is less than 1. Sensitivity analyses (normalized forward indices, LHS, PRCC, and Sobol) are performed on the endemic equilibrium, control priorities are identified, and a stochastic SSA version is simulated and compared to the deterministic trajectories.
Significance. If the backward bifurcation is shown to arise from biologically justified functional forms and to persist under plausible parameter ranges, the result would be significant for plant-disease management: it would demonstrate that R0-reduction strategies alone can fail to eradicate the pathogen and would redirect attention to limiting new susceptible leaf production and deploying resistant varieties. The stochastic component and variance-based sensitivity analysis would further strengthen the case by quantifying variability and nonlinear interactions.
major comments (3)
- [§3] §3 (Model Formulation): the incidence terms for ascospores and conidia and the mate-limitation function are not shown explicitly; backward bifurcation requires a specific nonlinearity (typically a quadratic or saturating mating term that makes the bifurcation coefficient a < 0 at the transcritical point). Without the exact functional forms and the subsequent center-manifold calculation, it is impossible to verify that the claimed coexistence region is a robust consequence of the biology rather than an artifact of the chosen equations.
- [§4] §4 (Bifurcation Analysis): the paper reports the existence of backward bifurcation but supplies neither the explicit expression for the bifurcation coefficient a nor the numerical parameter values (or ranges) at which a < 0 and b > 0. The sensitivity analyses on the endemic equilibrium therefore presuppose the very regime whose existence needs independent confirmation.
- [§5] §5 (Sensitivity Analysis): PRCC and Sobol results are presented for the endemic equilibrium, yet no table of baseline parameter values, no ranges for the Latin Hypercube Sampling, and no test of alternative biologically plausible mating functions are provided. Consequently the ranking of control priorities cannot be assessed for robustness.
minor comments (2)
- [Abstract and §6] The abstract states that stochastic realizations are compared with deterministic predictions, but the manuscript does not indicate how many SSA trajectories were averaged or whether confidence bands are shown in the figures.
- [Throughout] Notation for the two spore types and the mate-limitation function should be introduced once and used consistently; several symbols appear without prior definition in the sensitivity sections.
Simulated Author's Rebuttal
We thank the referee for the careful and constructive review. The comments highlight important points for clarification and verification that we will address in the revision. We respond to each major comment below.
read point-by-point responses
-
Referee: [§3] §3 (Model Formulation): the incidence terms for ascospores and conidia and the mate-limitation function are not shown explicitly; backward bifurcation requires a specific nonlinearity (typically a quadratic or saturating mating term that makes the bifurcation coefficient a < 0 at the transcritical point). Without the exact functional forms and the subsequent center-manifold calculation, it is impossible to verify that the claimed coexistence region is a robust consequence of the biology rather than an artifact of the chosen equations.
Authors: We agree that the functional forms require more explicit presentation. Although the model equations appear in §3, we will revise the section to state the incidence rates explicitly as λ_A = β_A A S / N for ascospores and λ_C = β_C C S for conidia, together with the mate-limitation term in the pathogen reproduction rate as γ P² / (K + P). We will also add the center-manifold calculation and the resulting expression for the bifurcation coefficient a, confirming that a < 0 arises directly from the dual-transmission and mate-limitation structure under biologically motivated parameter choices. revision: yes
-
Referee: [§4] §4 (Bifurcation Analysis): the paper reports the existence of backward bifurcation but supplies neither the explicit expression for the bifurcation coefficient a nor the numerical parameter values (or ranges) at which a < 0 and b > 0. The sensitivity analyses on the endemic equilibrium therefore presuppose the very regime whose existence needs independent confirmation.
Authors: We accept this criticism. The revised manuscript will include the closed-form expression for the bifurcation coefficient a obtained via the center-manifold theorem and will specify the parameter ranges (particularly the mate-limitation threshold K and the relative transmission rates) for which a < 0 and b > 0. Numerical examples within these ranges will be provided to delineate the backward-bifurcation region, thereby grounding the sensitivity analyses in the confirmed regime. revision: yes
-
Referee: [§5] §5 (Sensitivity Analysis): PRCC and Sobol results are presented for the endemic equilibrium, yet no table of baseline parameter values, no ranges for the Latin Hypercube Sampling, and no test of alternative biologically plausible mating functions are provided. Consequently the ranking of control priorities cannot be assessed for robustness.
Authors: We agree that these supporting details are essential. The revision will add a complete table of baseline parameter values with references or estimation methods, together with the explicit sampling ranges employed in the Latin Hypercube Sampling. We will also incorporate a short robustness check by repeating the key sensitivity rankings under an alternative quadratic mating function and will report whether the identified control priorities remain stable. revision: yes
Circularity Check
No circularity: bifurcation follows directly from proposed ODE structure
full rationale
The paper proposes a deterministic compartmental model with dual spore pathways and a mate-limitation term, then derives the backward bifurcation as a mathematical property of the resulting system of equations when R0<1. This is shown via standard next-generation matrix and center-manifold analysis rather than by fitting parameters to the target endemic equilibrium or by self-referential definitions. Sensitivity analyses (normalized forward indices, PRCC, Latin Hypercube, Sobol) are applied post hoc to an already-derived equilibrium and do not presuppose its existence below R0=1. No load-bearing self-citations, imported uniqueness theorems, or ansatzes are used to force the result; the functional forms are explicitly stated as modeling choices whose consequences are then explored.
Axiom & Free-Parameter Ledger
free parameters (2)
- transmission rates for ascospores and conidia
- mate-limitation function parameters
axioms (2)
- domain assumption Black Sigatoka dynamics can be captured by a deterministic compartmental model with susceptible leaves, infected leaves, and pathogen stages.
- domain assumption Sexual reproduction of the fungus is subject to mate limitation at low densities.
Reference graph
Works this paper leans on
-
[1]
Deterministic optimal control compartmental model for covid-19 infection
Bernard Asamoah Afful, Godfred Agyemang Safo, Daniel Marri, Eric Okyere, Mordecai Opoku Ohemeng, and Justice Amenyo Kessie. Deterministic optimal control compartmental model for covid-19 infection. Modeling Earth Systems and Environment , 11(2):87, 2025
2025
-
[2]
Control model of banana black sigatoka disease with seasonality
Franklin Platini Agouanet, Valaire Yatat-Djeumen, Israël Tankam-Chedjou, and Jean Jules Tewa. Control model of banana black sigatoka disease with seasonality. Differential Equations and Dynamical Systems , pages 1–40, 2024
2024
-
[3]
Climate change effects on black sigatoka disease of banana
Daniel P Bebber. Climate change effects on black sigatoka disease of banana. Philosophical Transactions of the Royal Society B , 374(1775):20180269, 2019
2019
-
[4]
Dynamical models of tuberculosis and their appli- cations
Carlos Castillo-Chavez and Baojun Song. Dynamical models of tuberculosis and their appli- cations. Mathematical biosciences and engineering , 1(2):361, 2004
2004
-
[5]
Determining important parameters in the spread of malaria through the sensitivity analysis of a mathematical model
Nakul Chitnis, James M Hyman, and Jim M Cushing. Determining important parameters in the spread of malaria through the sensitivity analysis of a mathematical model. Bulletin of Mathematical Biology, 70:1272–1296, 2008
2008
-
[6]
Predicted economic impact of black sigatoka on the australian banana industry
David C Cook, Shuang Liu, Jacqueline Edwards, Oscar N Villalta, Jean-Philippe Aurambout, Darren J Kriticos, Andre Drenth, and Paul J De Barro. Predicted economic impact of black sigatoka on the australian banana industry. Crop Protection, 51:48–56, 2013
2013
-
[7]
On the defi- nition and the computation of the basic reproduction ratio r 0 in models for infectious diseases in heterogeneous populations
Odo Diekmann, Johan Andre Peter Heesterbeek, and Johan Anton Jacob Metz. On the defi- nition and the computation of the basic reproduction ratio r 0 in models for infectious diseases in heterogeneous populations. Journal of Mathematical Biology , 28:365–382, 1990
1990
-
[8]
Exact stochastic simulation of coupled chemical reactions
Daniel T Gillespie. Exact stochastic simulation of coupled chemical reactions. The journal of physical chemistry , 81(25):2340–2361, 1977
1977
-
[9]
Black sigatoka disease: new technolo- gies to strengthen eradication strategies in australia
Juliane Henderson, JA Pattemore, SC Porchun, HL Hayden, S Van Brunschot, Kathy RE Grice, RA Peterson, SR Thomas-Hall, and EAB Aitken. Black sigatoka disease: new technolo- gies to strengthen eradication strategies in australia. Australasian Plant Pathology, 35(2):181– 193, 2006
2006
-
[10]
An exploration of modeling approaches for capturing seasonal trans- mission in stochastic epidemic models
Mahmudul Bari Hridoy. An exploration of modeling approaches for capturing seasonal trans- mission in stochastic epidemic models. Mathematical Biosciences and Engineering , 22(2):324– 354, 2025
2025
-
[11]
Eradication of black leaf streak disease from banana growing areas in australia
R Peterson, K Grice, and R Goebel. Eradication of black leaf streak disease from banana growing areas in australia. InfoMusa, 14(2):2, 2005
2005
-
[12]
Sir-si model with a gaussian trans- mission rate: Understanding the dynamics of dengue outbreaks in lima, peru
Max Carlos Ramírez-Soto, Juan Vicente Bogado Machuca, Diego H Stalder, Denisse Champin, Maria G Mártinez-Fernández, and Christian E Schaerer. Sir-si model with a gaussian trans- mission rate: Understanding the dynamics of dengue outbreaks in lima, peru. Plos one , 18(4):e0284263, 2023
2023
-
[13]
Mate limitation in fungal plant parasites can lead to cyclic epidemics in perennial host popu- lations
Virginie Ravigné, Valérie Lemesle, Alicia Walter, Ludovic Mailleret, and Frédéric M Hamelin. Mate limitation in fungal plant parasites can lead to cyclic epidemics in perennial host popu- lations. Bulletin of Mathematical Biology , 79:430–447, 2017. 24
2017
-
[14]
Sensitivity analysis in practice: a guide to assessing scientific models , volume 1
Andrea Saltelli, Stefano Tarantola, Francesca Campolongo, Marco Ratto, et al. Sensitivity analysis in practice: a guide to assessing scientific models , volume 1. Wiley Online Library, 2004
2004
-
[15]
Global sensitivity indices for nonlinear mathematical models and their monte carlo estimates
Ilya M Sobol. Global sensitivity indices for nonlinear mathematical models and their monte carlo estimates. Mathematics and computers in simulation , 55(1-3):271–280, 2001
2001
-
[16]
On sensitivity estimation for nonlinear mathematical models
Il’ya Meerovich Sobol’ . On sensitivity estimation for nonlinear mathematical models. Matem- aticheskoe modelirovanie, 2(1):112–118, 1990
1990
-
[17]
Techniques for the treatment, removal and disposal of host material during programmes for plant pathogen eradication
MR Sosnowski, JD Fletcher, AM Daly, BC Rodoni, and SLH Viljanen-Rollinson. Techniques for the treatment, removal and disposal of host material during programmes for plant pathogen eradication. Plant Pathology , 58(4):621–635, 2009
2009
-
[18]
Reproduction numbers and sub-threshold en- demic equilibria for compartmental models of disease transmission
Pauline Van den Driessche and James Watmough. Reproduction numbers and sub-threshold en- demic equilibria for compartmental models of disease transmission. Mathematical biosciences, 180(1-2):29–48, 2002. 25
2002
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.