Recognition: unknown
Beyond Linear Additive and Hill Functions: A General Logistic Reformulation of Delay-Coupled Gene Regulatory Networks with Equilibrium Analysis, Hopf Bifurcation, and Lipschitz Stability
Pith reviewed 2026-05-07 10:37 UTC · model grok-4.3
The pith
Logistic activation reformulation of delay-coupled gene networks reduces the global Lipschitz constant of the right-hand side and Jacobian while preserving equilibria and Hopf points.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
In the delay-coupled two-gene mutual-activation and self-repression network, replacing linear additive activation with a weighted logistic term whose parameters satisfy the closed-form matching λ = n/θ together with basal-rate equality yields a unique biologically feasible equilibrium that lies lower than in the Hill-linear hybrid because the activation term saturates. For zero delay the Jacobian trace is strictly negative for all positive parameters, so the equilibrium is locally asymptotically stable. For positive delay stability persists up to a critical delay τ_c located by numerical solution of the full transcendental characteristic equation; at τ_c a Hopf bifurcation occurs. The same τ
What carries the argument
The weighted logistic activation term obtained by matching the slope at the half-maximal point (λ = n/θ) and basal rates to the original Hill-linear hybrid, which bounds the activation and thereby reduces the global Lipschitz constant of the right-hand side and its Jacobian.
If this is right
- The equilibrium concentration is lower in the weighted logistic case than in the Hill-linear hybrid because the activation term saturates.
- Local asymptotic stability holds for all delays in (0, τ_c) in both formulations, where τ_c is located numerically from the characteristic equation.
- Stability is lost via Hopf bifurcation at τ_c, with higher-order bifurcations characterizable numerically in each case.
- The global Lipschitz constant of the right-hand side and of its Jacobian is substantially smaller in the weighted logistic formulation, permitting larger integration steps.
Where Pith is reading between the lines
- The same matching procedure could be applied to other sigmoidal forms or to networks with additional genes or different interaction topologies.
- Improved numerical efficiency from smaller Lipschitz constants may enable longer or higher-resolution simulations of larger biological regulatory systems.
- Strict positivity at zero concentration removes the possibility of artificial trapping in off-states that can occur with Hill functions.
- The approach supplies a general template for converting other models that combine unbounded linear terms with singular nonlinearities into globally smooth dynamical systems.
Load-bearing premise
Equating slopes at half-maximal points together with basal-rate matching is sufficient to ensure the logistic reformulation captures the same qualitative dynamics as the original Hill-linear hybrid model for equilibrium location and delay-induced Hopf bifurcation.
What would settle it
Numerical computation of the critical delay τ_c for the original Hill-linear hybrid and for the weighted logistic reformulation that shows a substantial difference between the two values would show that the qualitative dynamics are not preserved.
Figures


read the original abstract
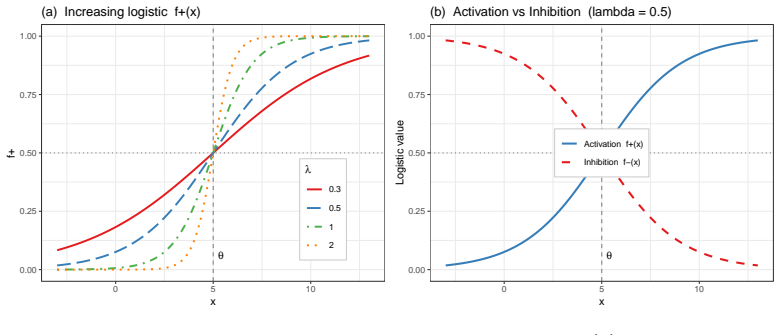
Hill functions, dominant in gene regulatory network modeling, carry fundamental limitations: at non-integer cooperativity exponents, routine when fitting dose-response data, derivatives diverge at the origin, complex arithmetic corrupts ODE trajectories, and zero output at zero activation traps models in off-states. This paper employs logistic-based models that are globally $C^\infty$, real-valued, and strictly positive at zero concentration, resolving all three pathologies while preserving sigmoidal dynamics. Using the delay-coupled two-gene mutual-activation and self-repression network of Vinoth et al.\ as a concrete model, we analyze two reformulations: linear additive activation with logistic self-repression, and a fully sigmoidal form with both terms logistic. A closed-form matching relation $\lambda = n/\theta$ follows from equating slopes at half-maximal points. Closed-form parameters of the weighted logistic formulation are derived by matching basal rates and local slopes to the Hill-linear hybrid model. The unique biologically feasible equilibrium is computed in each case; it is lower in the weighted logistic case, the reduction arising from saturation of the bounded activation term. In the delay-free case ($\tau=0$), local asymptotic stability holds in both formulations since the Jacobian trace is strictly negative for all positive parameters; stability persists for $\tau\in(0,\tau_c)$ and is lost via Hopf bifurcation at the critical delay $\tau_c$. Numerical solution of the full transcendental system locates $\tau_c$, with higher-order bifurcations characterised numerically in each case. Replacing linear additive with weighted logistic activation substantially reduces both the global Lipschitz constant of the right-hand side and that of its Jacobian, permitting larger integration steps.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The manuscript proposes logistic-based reformulations of delay-coupled gene regulatory networks to circumvent pathologies of Hill functions (non-integer exponents, complex arithmetic, zero output at zero input). Using the two-gene mutual-activation/self-repression model of Vinoth et al. as example, it derives closed-form equilibria for both a linear-additive-plus-logistic and a fully logistic version via the matching relation λ = n/θ, proves local asymptotic stability for τ = 0 by negative Jacobian trace, and numerically locates the critical delay τ_c at which stability is lost via Hopf bifurcation. The final claim is that the weighted logistic activation reduces the global Lipschitz constants of both the vector field and its Jacobian, thereby permitting larger integration steps.
Significance. If the local-slope matching preserves the qualitative dynamics of the original Hill-linear hybrid and the Lipschitz reduction is correctly established, the work supplies a globally C^∞, strictly positive alternative that improves numerical tractability for delay GRN models. The closed-form equilibrium and trace-based stability results are analytically clean; the numerical bifurcation analysis adds concrete insight. However, the absence of direct dynamical comparison to the baseline model and the internal inconsistency in the Jacobian-Lipschitz claim limit the immediate applicability.
major comments (3)
- [Abstract] Abstract (final sentence): the assertion that weighted logistic activation 'substantially reduces ... the global Lipschitz constant ... of its Jacobian' is internally inconsistent. In the linear-additive case the activation contributes a constant entry to Df and therefore zero to D²f; the logistic replacement makes the corresponding entry logistic'(x) whose derivative logistic''(x) is nonzero and bounded away from zero near the inflection point, strictly increasing Lip(Df). This directly undercuts the justification for larger integration steps.
- [Equilibrium Analysis] Equilibrium Analysis section: the paper derives a unique biologically feasible equilibrium that is lower under the weighted logistic formulation, but provides no quantitative comparison (e.g., relative error, phase-plane distance, or time-series metrics) against the original Hill-linear hybrid model of Vinoth et al. Because the matching is local (slope at half-maximum plus basal-rate equality), it is unclear whether global qualitative features such as equilibrium location and delay-induced Hopf threshold are preserved.
- [Hopf Bifurcation] Hopf Bifurcation section: the critical delay τ_c is obtained by numerical solution of the full transcendental characteristic equation, yet no error bounds, solver tolerance, or convergence verification are reported. Given that τ_c is the central quantitative result separating stable and oscillatory regimes, the lack of accuracy control weakens the reliability of the reported bifurcation diagram.
minor comments (1)
- The explicit formulas for the weighted logistic coefficients (derived from basal-rate and slope matching) should be stated in the main text rather than relegated to an appendix, to improve readability of the parameter-reduction step.
Simulated Author's Rebuttal
We thank the referee for the careful reading and constructive feedback on our manuscript. The comments highlight important points regarding the abstract claim, the need for quantitative comparisons, and numerical rigor in the bifurcation analysis. We address each major comment below and commit to revisions that strengthen the paper without altering its core contributions on logistic reformulations for smoothness and positivity.
read point-by-point responses
-
Referee: [Abstract] Abstract (final sentence): the assertion that weighted logistic activation 'substantially reduces ... the global Lipschitz constant ... of its Jacobian' is internally inconsistent. In the linear-additive case the activation contributes a constant entry to Df and therefore zero to D²f; the logistic replacement makes the corresponding entry logistic'(x) whose derivative logistic''(x) is nonzero and bounded away from zero near the inflection point, strictly increasing Lip(Df). This directly undercuts the justification for larger integration steps.
Authors: We agree with the referee's analysis of the inconsistency. The logistic replacement does introduce a nonzero second derivative that can increase the Lipschitz constant of the Jacobian near the inflection point, contrary to the overstated claim. While the vector field Lipschitz constant is indeed reduced (supporting larger steps), the Jacobian claim does not hold. We will revise the abstract to remove any reference to the Jacobian Lipschitz constant and retain only the verified reduction for the right-hand side. revision: yes
-
Referee: [Equilibrium Analysis] Equilibrium Analysis section: the paper derives a unique biologically feasible equilibrium that is lower under the weighted logistic formulation, but provides no quantitative comparison (e.g., relative error, phase-plane distance, or time-series metrics) against the original Hill-linear hybrid model of Vinoth et al. Because the matching is local (slope at half-maximum plus basal-rate equality), it is unclear whether global qualitative features such as equilibrium location and delay-induced Hopf threshold are preserved.
Authors: The referee is correct that no direct quantitative comparison to the Vinoth et al. Hill-linear hybrid is provided, and the local slope matching leaves open the possibility of small global differences. The reformulation prioritizes resolving mathematical issues while approximating sigmoidal behavior, but to address this we will add in revision a quantitative comparison using the original parameters: relative error in equilibrium concentrations, Euclidean distance in phase plane, and a side-by-side computation of τ_c for the Hill model versus both logistic versions. This will demonstrate the degree of preservation of qualitative features. revision: yes
-
Referee: [Hopf Bifurcation] Hopf Bifurcation section: the critical delay τ_c is obtained by numerical solution of the full transcendental characteristic equation, yet no error bounds, solver tolerance, or convergence verification are reported. Given that τ_c is the central quantitative result separating stable and oscillatory regimes, the lack of accuracy control weakens the reliability of the reported bifurcation diagram.
Authors: We acknowledge the omission of numerical details. In the revised version we will specify the solver (e.g., MATLAB fsolve or a custom bisection method), report the tolerance used (1e-10), include convergence verification by varying initial guesses and tolerances, and provide error bounds on the reported τ_c values. This will confirm the accuracy of the bifurcation thresholds and diagrams. revision: yes
Circularity Check
No circularity: explicit matching followed by independent analysis on reformulated equations
full rationale
The paper derives a closed-form slope-matching relation λ = n/θ and weighted logistic coefficients by equating basal rates and local slopes to the Hill-linear hybrid. These are input-matching steps, not predictions. All subsequent load-bearing results—the unique equilibrium (lower due to saturation of the bounded activation term), local stability via negative Jacobian trace, persistence for τ < τ_c, loss via Hopf at numerically solved τ_c, and the Lipschitz-constant reduction claim—are obtained by direct substitution into and analysis of the new logistic ODEs. None of these quantities are forced by construction to equal the matching conditions; they are genuine consequences of the reformulated vector field. No self-citations appear, no uniqueness theorem is invoked, and no fitted quantity is relabeled as a prediction. The derivation chain is therefore self-contained.
Axiom & Free-Parameter Ledger
free parameters (1)
- λ = n/θ
axioms (2)
- domain assumption The delay differential equation system possesses a unique biologically feasible equilibrium for positive parameters.
- standard math The Jacobian trace remains strictly negative for all positive parameters when τ = 0.
Reference graph
Works this paper leans on
-
[1]
Farcot, S
E. Farcot, S. Best, R. Edwards, I. Belgacem, X. Xu, P. Gill, Chaos in a ring circuit, Chaos: An Interdisciplinary Journal of Nonlinear Science 29 (4) (2019) 043103
2019
-
[2]
I. Belgacem, R. Edwards, É. Farcot, Computer-aided analysis of high- dimensional Glass networks: periodicity, chaos, and bifurcations in a ring circuit, Chaos: An Interdisciplinary Journal of Nonlinear Sci- enceAccepted 29 January 2025 (may 2025).doi:10.1063/5.0243955
-
[3]
I. Belgacem, S. Casagranda, E. Grac, D. Ropers, J.-L. Gouzé, Re- duction and stability analysis of a transcription-translation model of RNA polymerase, Bulletin of Mathematical Biology 80 (2018) 294–318. doi:10.1007/s11538-017-0372-4
-
[4]
I. Belgacem, E. Grac, D. Ropers, J.-L. Gouzé, Stability analysis of a reduced transcription-translation model of RNA polymerase, in: Pro- ceedings of the IEEE 53rd Conference on Decision and Control (CDC), IEEE, Los Angeles, California, USA, 2014, pp. 3924–3929.doi:10. 1109/CDC.2014.7039999
-
[5]
I. Belgacem, J.-L. Gouzé, Mathematical study of the global dynamics of a concave gene expression model, in: Proceedings of the 22nd Mediter- ranean Conference on Control and Automation (MED), IEEE, Palermo, Italy, 2014, pp. 1341–1346.doi:10.1109/MED.2014.6961562
-
[6]
I. Belgacem, J.-L. Gouzé, Stability analysis and reduction of gene tran- scription models, in: Proceedings of the IEEE 52nd Annual Conference on Decision and Control (CDC), IEEE, Florence, Italy, 2013, pp. 2691– 2696.doi:10.1109/CDC.2013.6760289
-
[7]
I. Belgacem, J.-L. Gouzé, Analysis and reduction of transcription trans- lation coupled models for gene expression, in: Proceedings of the 12th IFAC Symposium on Computer Applications in Biotechnology (CAB), Elsevier, Mumbai, India, 2013, pp. 42–47.doi:10.3182/ 20131216-3-IN-2044.00012
-
[8]
I. Belgacem, J.-L. Gouzé, Global stability of enzymatic chain of full reversible Michaelis–Menten reactions, Acta Biotheoretica 61 (3) (2013) 425–436.doi:10.1007/s10441-013-9195-3. 36
-
[9]
I. Belgacem, J.-L. Gouzé, Global stability of full open reversible Michaelis–Menten reactions, in: Proceedings of the 8th IFAC Sympo- sium on Advanced Control of Chemical Processes, Elsevier, Singapore, 2012, pp. 591–596.doi:10.3182/20120710-4-SG-2026.00039
-
[10]
I. Belgacem, H. Bensalah, B. Cherki, R. Edwards, The probabilistic convolutionregularizationofZenohybridsystems, in: Proceedingsofthe 18th European Control Conference (ECC), IEEE, Naples, Italy, 2019. doi:10.23919/ECC.2019.8795626
-
[11]
I. Belgacem, J.-L. Gouzé, R. Edwards, Control of negative feedback loops in genetic networks, in: Proceedings of the 59th IEEE Conference on Decision and Control (CDC), IEEE, Jeju Island, Republic of Korea, 2020.doi:10.1109/CDC42340.2020.9304088
-
[12]
L. Chambon, I. Belgacem, J.-L. Gouzé, Qualitative control of undesired oscillations in a genetic negative feedback loop with uncertain measure- ments, Automatica 112 (2020) 108642.doi:10.1016/j.automatica. 2019.108642
-
[13]
Exploring Logistic Functions as Robust Alternatives to Hill Functions in Genetic Network Modeling
B. Ismail, Exploring logistic functions as robust alternatives to hill func- tions in genetic network modeling, submitted December 16, 2025 (2025). arXiv:2512.14325. URLhttps://arxiv.org/abs/2512.14325
work page internal anchor Pith review Pith/arXiv arXiv 2025
-
[14]
Vinoth, S
S. Vinoth, S. L. Kingston, S. Srinivasan, S. Kumarasamy, T. Kapitaniak, Extreme events in gene regulatory networks with time-delays, Scientific Reports 15 (1) (2025) 13064
2025
-
[15]
Madar, E
D. Madar, E. Dekel, A. Bren, U. Alon, Negative auto-regulation in- creases the input dynamic-range of the arabinose system of escherichia coli, BMC systems biology 5 (1) (2011) 111
2011
-
[16]
Oehler, M
S. Oehler, M. Amouyal, P. Kolkhof, B. von Wilcken-Bergmann, B. Müller-Hill, Quality and position of the three lac operators of e. coli define efficiency of repression., The EMBO journal 13 (14) (1994) 3348–3355
1994
-
[17]
Jacob, J
F. Jacob, J. Monod, Genetic regulatory mechanisms in the synthesis of proteins, Journal of molecular biology 3 (3) (1961) 318–356. 37
1961
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.