Exploring Logistic Functions as Robust Alternatives to Hill Functions in Genetic Network Modeling
Pith reviewed 2026-05-16 22:12 UTC · model grok-4.3
The pith
Slope-matched logistic functions replace Hill functions in gene regulatory networks, eliminating non-integer coefficient pathologies and guaranteeing globally unique smooth bounded solutions.
A machine-rendered reading of the paper's core claim, the machinery that carries it, and where it could break.
Core claim
The product-of-logistics GRN model admits globally unique, smooth, uniformly bounded solutions with explicit Lipschitz constant L_F ≤ M = max_i(κ_i ∑_j L_i^j + γ_i). Each of the four Hill pathologies for non-integer n is resolved by a corresponding structural property of the logistic: the uniform bound |∂f^±/∂x| ≤ λ/4, the closed-form logit inverse, an elementary antiderivative, and the strictly positive basal output f^+(0) > 0. Although the Hill activation equals the logistic of the log-ratio, the change of variable s = ln(x/θ) multiplies the production term by a state-dependent factor e^{-s}, rendering the two ODE systems nonequivalent.
What carries the argument
The slope-matched logistic replacement λ = n/θ that supplies the uniform derivative bound |∂f/∂x| ≤ λ/4, making the vector field of the product-of-logistics ODE globally Lipschitz.
If this is right
- Standard contraction-mapping or Lyapunov arguments become directly applicable because the Lipschitz constant is known explicitly.
- Inverse problems for parameter estimation are well-posed since the logit inverse is single-valued and elementary.
- Numerical simulation requires no special handling for fractional exponents or singular derivatives.
- Basal expression remains strictly positive even at zero input, removing the artificial zero-production point of the Hill form.
Where Pith is reading between the lines
- Empirical model selection should compare logistic and Hill versions on the same data rather than defaulting to Hill, because the two encode different sensitivity hypotheses.
- Stochastic versions of the same GRN would inherit the same Lipschitz bound and therefore admit unique strong solutions.
- The strictly positive basal output may produce measurable leakage in long-term simulations that pure Hill models suppress to exact zero.
Load-bearing premise
That matching the derivative only at the half-maximal concentration is enough to make the logistic a biologically faithful stand-in for the Hill function across the whole range of concentrations.
What would settle it
Numerical integration of both models from the same initial conditions with identical parameters, followed by comparison of their trajectories against the same experimental time-series; systematic deviation in either transient shape or steady-state levels that cannot be removed by re-fitting parameters would falsify the replacement claim.
Figures








read the original abstract
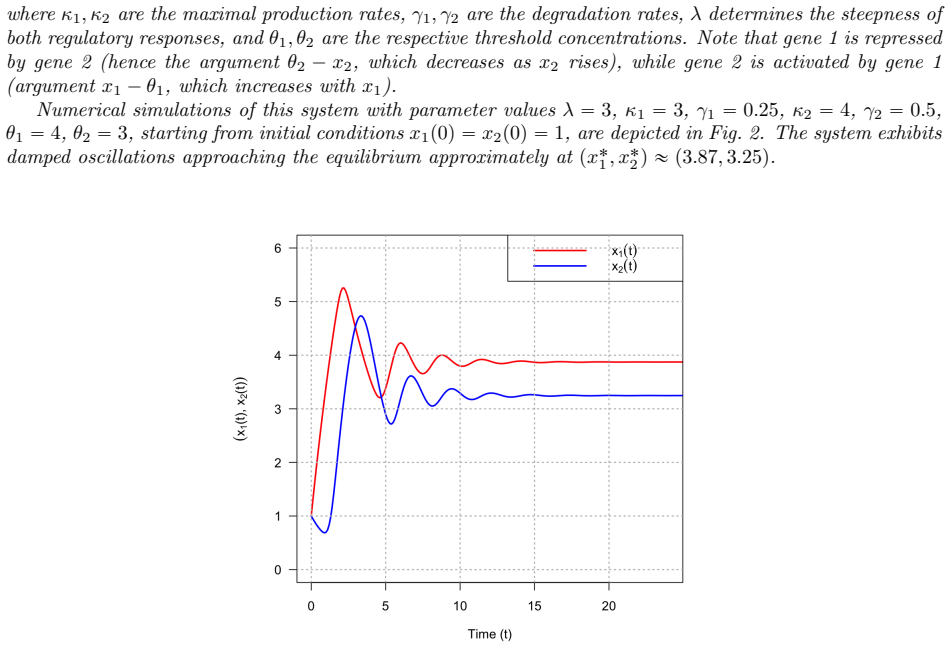
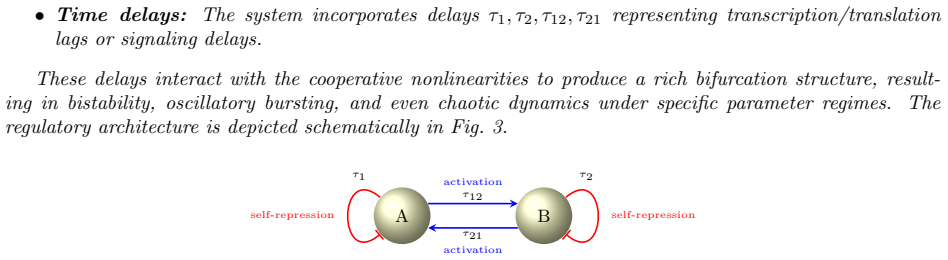
Hill functions dominate gene regulatory network (GRN) modeling, but their fractional exponents create analytical pathologies when the Hill coefficient $n$ is non-integer -- a ubiquitous occurrence in experimental fits. We replace the Hill activation $h^+(x,\theta,n)=x^n/(x^n+\theta^n)$ and repression $h^-(x,\theta,n)=\theta^n/(x^n+\theta^n)$ with the logistic counterparts $f^+(x,\theta,\lambda)=1/(1+e^{-\lambda(x-\theta)})$ and $f^-(x,\theta,\lambda)=1/(1+e^{\lambda(x-\theta)})$. The matching $\lambda=n/\theta$ preserves the slope at the half-maximal concentration. Four families of Hill pathologies appear for non-integer $n$: derivative singularities at the origin ($h^{+\prime}(x)\to\infty$ as $x\to 0^+$ for $0<n<1$; higher-order derivatives diverging for $n\in(k,k+1)$); integrals requiring hypergeometric functions; multivalued fractional-power inversions; and logarithmic small-$n$ approximations diverging at low expression. Each is resolved by a structural property of the logistic: the uniform bound $|\partial f^\pm/\partial x|\le\lambda/4$, the closed-form logit inverse, an elementary antiderivative, and the nonzero basal output $f^+(0)=1/(1+e^{\lambda\theta})>0$. We prove the product-of-logistics GRN model admits globally unique, smooth, uniformly bounded solutions with explicit Lipschitz constant $L_F\le M=\max_i(\kappa_i\sum_j L_i^j+\gamma_i)$. The identity $h^+(x,\theta,n)=\sigma(n\ln(x/\theta))$ shows the Hill is a logistic of the log-ratio, but the change of variable $s=\ln(x/\theta)$ introduces a state-dependent factor $e^{-s}$ on the production side, so the two ODE models are nonequivalent. They encode different hypotheses -- multiplicative-increment versus additive-threshold sensitivity -- and the structural advantages of the logistic framework hold under either.
Editorial analysis
A structured set of objections, weighed in public.
Referee Report
Summary. The paper proposes logistic activation and repression functions as replacements for Hill functions in gene regulatory network (GRN) ODE models to eliminate analytical pathologies that arise for non-integer Hill coefficients. It matches slopes at the half-maximal point via λ = n/θ, shows that the logistic resolves derivative singularities, hypergeometric integrals, multivalued inverses, and divergent small-n approximations, establishes nonequivalence to the Hill model through the identity h^+(x,θ,n) = σ(n ln(x/θ)) and the resulting state-dependent factor, and proves global existence, uniqueness, smoothness, and uniform boundedness of solutions for the product-of-logistics system together with the explicit Lipschitz bound L_F ≤ M = max_i(κ_i ∑_j L_i^j + γ_i).
Significance. The global well-posedness result with an explicit, parameter-dependent Lipschitz constant is a concrete and useful contribution that supports rigorous stability and numerical analysis of GRN models. The structural resolution of the four listed Hill pathologies via uniform derivative bounds, closed-form inverses, and elementary antiderivatives is technically clean. The observation that the two modeling frameworks encode distinct hypotheses (multiplicative-increment versus additive-threshold sensitivity) is a helpful conceptual clarification.
major comments (1)
- Abstract (modeling paragraph): the assertion that λ = n/θ yields a 'biologically faithful replacement' is load-bearing for the paper's positioning of the logistic as a robust alternative, yet the manuscript provides no comparative dynamical simulations, steady-state analysis, or fitting to experimental GRN data to show that slope matching alone preserves response times, expression distributions, or network-level behavior; without such evidence the replacement claim remains mathematical rather than biologically substantiated.
minor comments (2)
- Abstract (pathology list): the four families of Hill pathologies are clearly enumerated, but the corresponding logistic resolutions would benefit from one-sentence pointers to the precise structural property (e.g., 'uniform bound |∂f/∂x| ≤ λ/4' for the derivative singularity) to improve readability.
- Proof of the Lipschitz bound (abstract and §3): the estimate |∏f_j(x) − ∏f_j(y)| ≤ ∑_j (λ_j/4) |x−y| is stated as immediate from the [0,1] range, but an explicit intermediate inequality showing how the product difference is controlled by the sum of individual differences would make the derivation fully self-contained.
Simulated Author's Rebuttal
We thank the referee for the positive assessment of the global well-posedness result and the structural resolution of the Hill pathologies. We address the single major comment below.
read point-by-point responses
-
Referee: Abstract (modeling paragraph): the assertion that λ = n/θ yields a 'biologically faithful replacement' is load-bearing for the paper's positioning of the logistic as a robust alternative, yet the manuscript provides no comparative dynamical simulations, steady-state analysis, or fitting to experimental GRN data to show that slope matching alone preserves response times, expression distributions, or network-level behavior; without such evidence the replacement claim remains mathematical rather than biologically substantiated.
Authors: We agree that the manuscript is a mathematical analysis and contains no comparative dynamical simulations, steady-state comparisons, or experimental data fitting. The slope-matching condition λ = n/θ is presented as a standard way to align the threshold sensitivity, but we acknowledge that this does not automatically guarantee preservation of network-level dynamics or biological equivalence. To avoid overstating the claim, we will revise the abstract (and the corresponding sentence in the introduction) to remove the phrase 'biologically faithful replacement' and instead state that the logistic provides a mathematically robust alternative that preserves the half-maximal slope while eliminating the listed analytical pathologies. The nonequivalence of the two modeling frameworks is already established in the manuscript via the state-dependent factor arising from the log-ratio change of variables; the revision will make this distinction more prominent in the abstract. revision: yes
Circularity Check
No significant circularity identified
full rationale
The paper derives global existence, uniqueness, smoothness, and uniform boundedness for the product-of-logistics GRN system directly from the intrinsic properties of the logistic functions (uniform derivative bound |∂f±/∂x| ≤ λ/4 and range [0,1]) together with the standard product Lipschitz estimate and linear degradation. These yield the explicit constant M without any parameter fitting, self-referential definitions, or load-bearing self-citations. The slope-matching choice λ = n/θ is presented as an explicit modeling assumption for local approximation, not as a derived or fitted prediction. The relation to Hill functions via logit is used only to contrast the models, not to justify the logistic results. The derivation is therefore self-contained against external mathematical benchmarks.
Axiom & Free-Parameter Ledger
axioms (1)
- standard math Picard-Lindelöf theorem: a Lipschitz-continuous right-hand side guarantees local existence and uniqueness of ODE solutions
Lean theorems connected to this paper
-
IndisputableMonolith/Cost/FunctionalEquation.leanwashburn_uniqueness_aczel unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
We replace the Hill activation h⁺(x,θ,n)=xⁿ/(xⁿ+θⁿ) and repression h⁻(x,θ,n)=θⁿ/(xⁿ+θⁿ) with the logistic counterparts f⁺(x,θ,λ)=1/(1+e^{-λ(x-θ)}) and f⁻(x,θ,λ)=1/(1+e^{λ(x-θ)}). The matching λ=n/θ preserves the slope at the half-maximal concentration. We prove the product-of-logistics GRN model admits globally unique, smooth, uniformly bounded solutions with explicit Lipschitz constant L_F ≤ M = max_i(κ_i ∑_j L_i^j + γ_i).
-
IndisputableMonolith/Foundation/ArithmeticFromLogic.leanembed_eq_pow unclear?
unclearRelation between the paper passage and the cited Recognition theorem.
The identity h⁺(x,θ,n)=σ(n ln(x/θ)) shows the Hill is a logistic of the log-ratio, but the change of variable s=ln(x/θ) introduces a state-dependent factor e^{-s} on the production side, so the two ODE models are nonequivalent.
What do these tags mean?
- matches
- The paper's claim is directly supported by a theorem in the formal canon.
- supports
- The theorem supports part of the paper's argument, but the paper may add assumptions or extra steps.
- extends
- The paper goes beyond the formal theorem; the theorem is a base layer rather than the whole result.
- uses
- The paper appears to rely on the theorem as machinery.
- contradicts
- The paper's claim conflicts with a theorem or certificate in the canon.
- unclear
- Pith found a possible connection, but the passage is too broad, indirect, or ambiguous to say the theorem truly supports the claim.
Forward citations
Cited by 2 Pith papers
-
Logistic Gene Regulatory Networks: Prevention of Expression Shutdown, and Numerical Stability Beyond Hill Function
Logistic functions replace Hill functions in gene regulatory network ODE models to ensure global smoothness, real-valued trajectories, positive basal production, and stable numerical integration across small and large...
-
Beyond Linear Additive and Hill Functions: A General Logistic Reformulation of Delay-Coupled Gene Regulatory Networks with Equilibrium Analysis, Hopf Bifurcation, and Lipschitz Stability
Logistic reformulations of delay-coupled gene regulatory networks are globally smooth and positive at zero, with matched parameters, unique equilibria, Hopf bifurcation at critical delays, and substantially smaller Li...
Reference graph
Works this paper leans on
-
[1]
I. Belgacem, S. Casagranda, E. Grac, D. Ropers, J.-L. Gouz´ e, Reduction and stability analysis of a transcription–translation model of rna polymerase, Bulletin of Mathematical Biology 80 (2) (2018) 294–318
work page 2018
-
[2]
I. Belgacem, J.-L. Gouz´ e, R. Edwards, Control of negative feedback loops in genetic networks, in: 2020 59th IEEE Conference on Decision and Control (CDC), IEEE, 2020, pp. 5098–5105
work page 2020
-
[3]
L. Chambon, I. Belgacem, J.-L. Gouz´ e, Qualitative control of undesired oscillations in a genetic negative feedback loop with uncertain measurements, Automatica 112 (2020) 108642
work page 2020
-
[4]
I. Belgacem, J.-L. Gouz´ e, Global stability of enzymatic chains of full reversible michaelis-menten reactions, Acta biotheoretica 61 (3) (2013) 425–436
work page 2013
-
[5]
G. Bernot, J.-P. Comet, A. Richard, M. Chaves, J.-L. Gouz´ e, F. Dayan, Modeling and analysis of gene regulatory networks, in: Modeling in computational biology and biomedicine: A multidisciplinary endeavor, Springer, 2012, pp. 47–80
work page 2012
-
[6]
W. Abou-Jaoud´ e, M. Chaves, J.-L. Gouz´ e, A theoretical exploration of birhythmicity in the p53-mdm2 network, PLOS one 6 (2) (2011) e17075
work page 2011
-
[7]
A. Polynikis, S. Hogan, M. di Bernardo, Comparing different ode modelling approaches for gene regu- latory networks, Journal of theoretical biology 261 (4) (2009) 511–530
work page 2009
-
[8]
S. Bottani, R. A. Veitia, Hill function-based models of transcriptional switches: impact of specific, nonspecific, functional and nonfunctional binding, Biological Reviews 92 (2) (2017) 953–963
work page 2017
-
[9]
H. Kim, E. Gelenbe, Stochastic gene expression modeling with hill function for switch-like gene re- sponses, IEEE/ACM Transactions on Computational Biology and Bioinformatics 9 (4) (2011) 973–979
work page 2011
-
[10]
M. Santill´ an, On the use of the hill functions in mathematical models of gene regulatory networks, Mathematical Modelling of Natural Phenomena 3 (2) (2008) 85–97
work page 2008
- [11]
-
[12]
H. De Jong, J.-L. Gouz´ e, C. Hernandez, M. Page, T. Sari, J. Geiselmann, Qualitative simulation of genetic regulatory networks using piecewise-linear models, Bulletin of mathematical biology 66 (2) (2004) 301–340
work page 2004
-
[13]
J.-L. Gouz´ e, T. Sari, A class of piecewise linear differential equations arising in biological models, Dynamical systems 17 (4) (2002) 299–316
work page 2002
- [14]
-
[15]
A. A. Nielsen, B. S. Der, J. Shin, P. Vaidyanathan, V. Paralanov, E. A. Strychalski, D. Ross, D. Dens- more, C. A. Voigt, Genetic circuit design automation, Science 352 (6281) (2016) aac7341
work page 2016
-
[16]
G. Rodrigo, A. Jaramillo, Autobiocad: full biodesign automation of genetic circuits, ACS synthetic biology 2 (5) (2013) 230–236
work page 2013
-
[17]
I. Otero-Muras, D. Henriques, J. R. Banga, Synbadm: a tool for optimization-based automated design of synthetic gene circuits, Bioinformatics 32 (21) (2016) 3360–3362
work page 2016
-
[18]
M. Stamatakis, N. V. Mantzaris, Comparison of deterministic and stochastic models of the lac operon genetic network, Biophysical journal 96 (3) (2009) 887–906. 81
work page 2009
-
[19]
E. M. Ozbudak, M. Thattai, H. N. Lim, B. I. Shraiman, A. Van Oudenaarden, Multistability in the lactose utilization network of escherichia coli, Nature 427 (6976) (2004) 737–740
work page 2004
-
[20]
M. Du, S. Kodner, L. Bai, Enhancement of laci binding in vivo, Nucleic acids research 47 (18) (2019) 9609–9618
work page 2019
-
[21]
U. Alon, An Introduction to Systems Biology: Design Principles of Biological Circuits, Chapman & Hall/CRC Mathematical and Computational Biology Series, Chapman & Hall/CRC, Boca Raton, FL, 2007, see Appendix A: The Input Functions of Genes: Michaelis–Menten and Hill Equations. URLhttps://www.weizmann.ac.il/mcb/UriAlon/sites/mcb.UriAlon/files/uploads/appen...
work page 2007
- [22]
-
[23]
T. S. Gardner, C. R. Cantor, J. J. Collins, Construction of a genetic toggle switch in escherichia coli, Nature 403 (6767) (2000) 339–342
work page 2000
-
[24]
M. B. Elowitz, S. Leibler, A synthetic oscillatory network of transcriptional regulators, Nature 403 (6767) (2000) 335
work page 2000
-
[25]
F. Farzadfard, T. K. Lu, Genomically encoded analog memory with precise in vivo dna writing in living cell populations, Science 346 (6211) (2014) 1256272
work page 2014
- [26]
-
[27]
A. Polynikis, S. J. Hogan, M. Di Bernardo, Comparing different ode modelling approaches for gene regulatory networks, Journal of theoretical biology 261 (4) (2009) 511–530
work page 2009
-
[28]
K. Hari, W. Duncan, M. A. Ibrahim, M. K. Jolly, B. Cummins, T. Gedeon, Assessing biological network dynamics: comparing numerical simulations with analytical decomposition of parameter space, npj Systems Biology and Applications 9 (1) (2023) 29
work page 2023
-
[29]
I. A. Mellis, M. E. Melzer, N. Bodkin, Y. Goyal, Prevalence of and gene regulatory constraints on transcriptional adaptation in single cells, Genome biology 25 (1) (2024) 217
work page 2024
-
[30]
F. M. Alakwaa, Modeling of gene regulatory networks: a literature review, Journal of Computational Systems Biology 1 (1) (2014) 1
work page 2014
-
[31]
P. Dibaeinia, S. Sinha, Sergio: a single-cell expression simulator guided by gene regulatory networks, Cell systems 11 (3) (2020) 252–271
work page 2020
-
[32]
H. Abeliovich, An empirical extremum principle for the hill coefficient in ligand-protein interactions showing negative cooperativity, Biophysical journal 89 (1) (2005) 76–79
work page 2005
- [33]
-
[34]
D. Leipply, D. E. Draper, Dependence of rna tertiary structural stability on mg2+ concentration: interpretation of the hill equation and coefficient, Biochemistry 49 (9) (2010) 1843–1853
work page 2010
-
[35]
G. Quee, R. Edwards, Ramp approximations of sigmoid control functions in gene networks, Physica D: Nonlinear Phenomena 418 (2021) 132840. 82
work page 2021
-
[36]
J. Berkson, A statistically precise and relatively simple method of estimating the bio-assay with quantal response, based on the logistic function, Journal of the American Statistical Association 48 (263) (1953) 565–599
work page 1953
-
[37]
Q. Ke, B. J. Oommen, Logistic neural networks: Their chaotic and pattern recognition properties, Neurocomputing 125 (2014) 184–194
work page 2014
-
[38]
P. E. Puddu, A. Menotti, Artificial neural network versus multiple logistic function to predict 25-year coronary heart disease mortality in the seven countries study, European Journal of Cardiovascular Prevention & Rehabilitation 16 (5) (2009) 583–591
work page 2009
-
[39]
S. Abolmaali, S. Shirzaei, A comparative study of sir model, linear regression, logistic function and arima model for forecasting covid-19 cases, AIMS public health 8 (4) (2021) 598
work page 2021
-
[40]
Y. Koizumi, T. Miyamura, S. Arakawa, E. Oki, K. Shiomoto, M. Murata, Adaptive virtual network topology control based on attractor selection, Journal of Lightwave Technology 28 (11) (2010) 1720– 1731
work page 2010
-
[41]
S. Goebbels, On sharpness of error bounds for univariate approximation by single hidden layer feed- forward neural networks, Results in Mathematics 75 (3) (2020) 109
work page 2020
-
[42]
I. Samuilik, F. Sadyrbaev, D. Ogorelova, Mathematical modeling of three-dimensional genetic regula- tory networks using logistic and gompertz functions, WSEAS Transactions on systems and control 17 (2022) 101107
work page 2022
-
[43]
F. Sadyrbaev, I. Samuilik, V. Sengileyev, On modelling of genetic regulatory networks, WSEAS Trans- actions on Electronics 12 (1) (2021) 73
work page 2021
-
[44]
O. Kozlovska, F. Sadyrbaev, In search of chaos in genetic systems, Chaos Theory and Applications 6 (1) (2024) 13–18
work page 2024
-
[45]
I. Samuilik, F. Sadyrbaev, Genetic engineering–construction of a network of arbitrary dimension with periodic attractor, Vibroengineering Procedia 46 (2022) 67–72
work page 2022
-
[46]
F. Sadyrbaev, V. Sengileyev, A. Silvans, On coexistence of inhibition and activation in genetic regu- latory networks, in: International Conference on Numerical Analysis and Applied Mathematics 2021, ICNAAM 2021, AIP PRESS, 2023
work page 2021
-
[47]
S. S. Somathilaka, S. Balasubramaniam, D. P. Martins, X. Li, Revealing gene regulation-based neural network computing in bacteria, Biophysical Reports 3 (3) (2023)
work page 2023
-
[48]
O. Kozlovska, F. Sadyrbaev, Modeling networks of four elements, Computation 13 (5) (2025) 123
work page 2025
-
[49]
T. W. Tee, A. Chowdhury, C. D. Maranas, J. V. Shanks, Systems metabolic engineering design: fatty acid production as an emerging case study, Biotechnology and bioengineering 111 (5) (2014) 849–857
work page 2014
-
[50]
T. Radivojevi´ c, Z. Costello, K. Workman, H. Garcia Martin, A machine learning automated recom- mendation tool for synthetic biology, Nature communications 11 (1) (2020) 4879
work page 2020
- [51]
-
[52]
L. Xie, W. Lu, J. Yu, Y. Zhang, H. Gao, C. Xie, Y. Tian, M. Liu, G. Wang, Regulation of insulin expression and release in gene and cell therapy of insulin-deficient diabetes, European Journal of Endocrinology (2025) lvaf231. 83
work page 2025
-
[53]
B. Xie, P. M. Nguyen, A. Guˇ cek, A. Thonig, S. Barg, O. Idevall-Hagren, Plasma membrane phos- phatidylinositol 4, 5-bisphosphate regulates ca2+-influx and insulin secretion from pancreaticβcells, Cell chemical biology 23 (7) (2016) 816–826
work page 2016
-
[54]
Z. Chen, F. Cao, The properties of logistic function and applications to neural network approximation, Journal of Computational Analysis and Applications 15 (6) (2013) 1046–1056
work page 2013
-
[55]
N. Kyurkchiev, S. Markov, Sigmoid functions: some approximation and modelling aspects, LAP LAM- BERT Academic Publishing, Saarbrucken 4 (2015) 34
work page 2015
-
[56]
B.-S. Chen, C.-W. Li, On the interplay between entropy and robustness of gene regulatory networks, Entropy 12 (5) (2010) 1071–1101
work page 2010
- [57]
- [58]
-
[59]
P. G. Gottschalk, J. R. Dunn, The five-parameter logistic: a characterization and comparison with the four-parameter logistic, Analytical biochemistry 343 (1) (2005) 54–65
work page 2005
-
[60]
R. Li, J. C. Rozum, M. M. Quail, M. N. Qasim, S. S. Sindi, C. J. Nobile, R. Albert, A. D. Hern- day, Inferring gene regulatory networks using transcriptional profiles as dynamical attractors, PLoS computational biology 19 (8) (2023) e1010991
work page 2023
-
[61]
M. Aguirre, J. P. Spence, G. Sella, J. K. Pritchard, Gene regulatory network structure informs the distribution of perturbation effects, PLOS Computational Biology 21 (9) (2025) e1013387
work page 2025
-
[62]
M. Abramowitz, I. A. Stegun, Handbook of Mathematical Functions with Formulas, Graphs, and Mathematical Tables, Dover Publications, 1965
work page 1965
-
[63]
Q. Yuan, Z. Duren, Inferring gene regulatory networks from single-cell multiome data using atlas-scale external data, Nature Biotechnology 43 (2) (2025) 247–257
work page 2025
-
[64]
J. Xu, C. Lu, S. Jin, Y. Meng, X. Fu, X. Zeng, R. Nussinov, F. Cheng, Deep learning-based cell-specific gene regulatory networks inferred from single-cell multiome data, Nucleic Acids Research 53 (5) (2025) gkaf138
work page 2025
- [65]
-
[66]
J. Kursawe, A. Moneyron, T. Galla, Efficient approximations of transcriptional bursting effects on the dynamics of a gene regulatory network, Journal of the Royal Society Interface 22 (227) (2025) 20250170
work page 2025
-
[67]
C. Chen, M. Padi, Flexible modeling of regulatory networks improves transcription factor activity estimation, NPJ Systems Biology and Applications 10 (1) (2024) 58
work page 2024
-
[68]
G. Mao, R. Zeng, J. Peng, K. Zuo, Z. Pang, J. Liu, Reconstructing gene regulatory networks of biological function using differential equations of multilayer perceptrons, BMC bioinformatics 23 (1) (2022) 503
work page 2022
-
[69]
S. A. Frank, Input-output relations in biological systems: measurement, information and the hill equation, Biology direct 8 (1) (2013) 31. 84
work page 2013
-
[70]
S. Naqvi, S. Kim, H. Hoskens, H. S. Matthews, R. A. Spritz, O. D. Klein, B. Hallgr´ ımsson, T. Swigut, P. Claes, J. K. Pritchard, et al., Precise modulation of transcription factor levels identifies features underlying dosage sensitivity, Nature genetics 55 (5) (2023) 841–851
work page 2023
-
[71]
M. Son, A. G. Wang, H.-L. Tu, M. O. Metzig, P. Patel, K. Husain, J. Lin, A. Murugan, A. Hoffmann, S. Tay, Nf-κb responds to absolute differences in cytokine concentrations, Science signaling 14 (666) (2021) eaaz4382
work page 2021
-
[72]
URLhttps://www.nature.com/articles/nature13182
The FANTOM Consortium and the RIKEN PMI and CLST (DGT), A promoter-level mammalian expression atlas, Nature 507 (7493) (2014) 462–470.doi:10.1038/nature13182. URLhttps://www.nature.com/articles/nature13182
- [73]
- [74]
- [75]
- [76]
-
[77]
O. Kozlovska, F. Sadyrbaev, Models of genetic networks with given properties, WSEAS Transactions on Computer Research 10 (2022) 43–49
work page 2022
-
[78]
R. Lutz, H. Bujard, Independent and tight regulation of transcriptional units in escherichia coli via the lacr/o, the tetr/o and arac/i1-i2 regulatory elements, Nucleic acids research 25 (6) (1997) 1203–1210
work page 1997
-
[79]
J. R. Kelly, A. J. Rubin, J. H. Davis, C. M. Ajo-Franklin, J. Cumbers, M. J. Czar, K. de Mora, A. L. Glieberman, D. D. Monie, D. Endy, Measuring the activity of biobrick promoters using an in vivo reference standard, Journal of biological engineering 3 (1) (2009) 4
work page 2009
-
[80]
I. Joanito, C.-C. S. Yan, J.-W. Chu, S.-H. Wu, C.-P. Hsu, Basal leakage in oscillation: Coupled transcriptional and translational control using feed-forward loops, PLOS Computational Biology 16 (9) (2020) e1007740
work page 2020
discussion (0)
Sign in with ORCID, Apple, or X to comment. Anyone can read and Pith papers without signing in.